ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 28.03.2024
Просмотров: 8
Скачиваний: 0
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.

Основы молекулярной генетики. Репликация. Транскрипция
Цель лекции
Ознакомиться с основными понятиями матричных процессов: репликации ДНК, транскрипции РНК, трансляции (биосинтеза белка), репарации ДНК. Уметь интерпретировать информацию о биосинтезе белка для объяснения адаптации организма к изменяющимся условиям среды и решения вопросов лечения и профилактики опухолевых и инфекционных болезней
План лекции
1. Понятие генетической информации и этапы ее передачи.
2. Биосинтез ДНК на ДНК-матрице (репликация). Основные этапы репликации.
3. Повреждения ДНК и их исправления (репарация)
4.Биосинтез РНК на ДНК-матрице (транскипция). Посттранскрипционные модификации РНК (процессинг)
1.Понятие генетической информации и этапы ее передачи
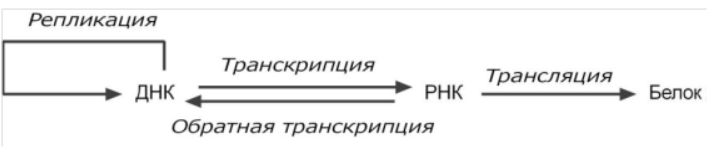
Нуклеиновые кислоты и белки представляют собой информационные молекулы. ДНК является хранителем генетической информации.
Поток информации передается от ДНК через РНК на белок.Этапы передачи генетической информации:
2.Биосинтез ДНК на ДНК-матрице (репликация)
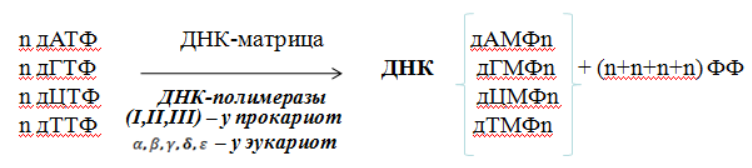
Репликация – биосинтез ДНК. Во время репликации каждая из двух цепей ДНК служит матрицей для образования новой цепи. Субстратами и источниками энергии для синтеза ДНК являются дезоксирибонуклеозидтрифосфаты (дНТФ: дАТФ, дГТФ, дЦТФ, дТТФ).
Суммарное уравнение репликации:
Процесс включает следующие основные этапы:
I. Формирование репликативной вилки (инициация).
II. Синтез новых цепей ДНК (элонгация).
III. Исключение праймеров. Завершение формирования отстающей цепи ДНК (терминация).
Биосинтез ДНК на ДНК-матрице (репликация). Основные этапы репликации.
2.1Инициация репликации
Синтез ДНК начинается в определенных участках, получивших название точка ori (англ. origin – начало). Происходит формирование «репликативной вилки» - места последовательного расплетения двойной цепи и синтеза новых цепей ДНК (рис. 1) при участии:
ДНК-топоизомеразы, которая является «обратимой нуклеазой». Фермент обеспечивает подготовку ДНК хромосом к репликации, разрывая 3'-,5'-фосфодиэфирную связь в цепи ДНК, способствуя ее раскручиванию;
ДНК-хеликаз - ДНК-зависимых АТФаз, использующих энергию АТФ для расплетения двойной спирали ДНК, путем разрушения водородных связей между комплементерными основаниями;
SSB (single strand binding)-белков, связывающихся с одноцепочечными участками ДНК. Эти белки, не закрывая оснований, предотвращают повторное комплементарное скручивание матричных цепей и образование шпилек.
2.2. Элонгация репликации
На этой стадии дочерние нити ДНК образуются на обеих нитях материнской ДНК. Процесс катализирует несколько ДНК-полимераз (I и III), которые синтезируют полинуклеотидные цепи из дНТФ: дАТФ, дГТФ, дТТФ и дЦТФ в направлении от 5'- к 3'-концу антипараллельно матрице, имеющей направление от 3'- к 5'-концу (рис. 1).
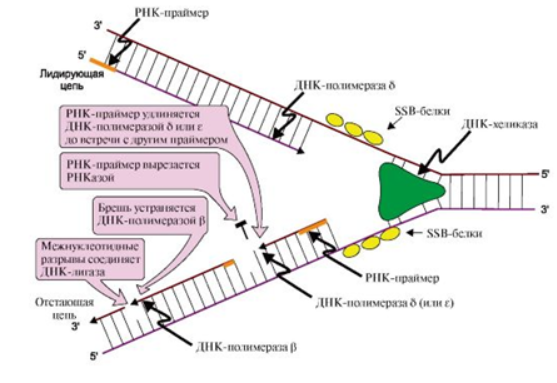
Рис. 1 Синтез новых цепей в области репликативной вилки
ДНК- полимераза I - расщепляет праймер (обладает нуклеазной активностью) и на его месте синтезирует комплиментарную цепь ДНК (обладает полимеразной активностью), заполняет бреши. ДНК-полимераза III - основной фермент репликации - синтезирует комплиментарные дочерние цепи на разошедшихся материнских цепях в направлении 5’→3’. Она не способна инициировать синтез новой цепи ДНК, может лишь удлинять затравку-праймер. Образует 3,5-фосфоэфирные связи между нуклеотидами, используя в качестве субстратов дезоксинуклеозидтрифосфаты (дНТФ).
Новые цепи синтезируются по-разному. На матрице ДНК с направлением от 3'- к 5'-концу цепь растет непрерывно по ходу движения репликативной вилки (→) и называется лидирующей. На матрице с направлением от 3'- к 5'-концу вторая цепь синтезируется против движения репликативной вилки (←) в виде коротких отрезков - фрагментов Оказаки. Рост этой цепи начинается только тогда, когда на матрице ДНК появляется одноцепочечный участок длиной около 200 нуклеотидов, поэтому ее называют запаздывающей или отстающей.
Лидирующая нить растет непрерывно, а отстающая - в виде фрагментов Оказаки, каждый из которых включает:
- РНК-праймер (
10 нуклеотидов);
- участок ДНК, примерно равный длине цепи из 150 нуклеотидов
ДНК-полимеразы I и III (δ, β и ε) не способны инициировать синтез новых цепей ДНК, они могут лишь удлинять имеющуюся нуклеотидную цепь. Синтез лидирующей и отстающей нитей начинается с образования затравки или праймера-олигорибонуклеотида (РНК) для ДНК-полимеразы III, включающего около 10 мононуклеотидов. Его образование катализируетпраймаза. Далее этот же фермент, используя в качестве субстратов дНТФ, переключается на образование ДНК и включает во вновь синтезируемую нить 20-50 дезоксирибонуклеотидов, после чего заменяется другими ДНК-полимеразами. Синтез лидирующей цепи продолжает ДНК-полимераза δ, а отстающей - ДНК-полимераза δ или ε. Оба фермента, помимо, полимеразной обладают еще и экзонуклеазной активностью. В ходе синтеза они могут исправлять допущенную ошибку и отщеплять неправильно включенный нуклеотид, что обеспечивает высокую точность синтеза ДНК.
2.3. Терминация репликации
Вырезание праймеров. Завершение формирования отстающей цепи ДНК. В отстающей нити праймер удаляется эндонуклеазой или РНКазой. Затем ДНК-полимераза I (ДНК-полимераза β) заполняет образованную «брешь», присоединяя по принципу комплементарности матрице дезоксирибонуклеотиды в количестве, равном вырезанному праймеру. ДНК-лигаза сшивает фрагменты синтезированной дочерней ДНК. Фермент, используя энергию АТФ, из множества фрагментов Оказаки образует непрерывную цепь ДНК. Результатом процесса является образование дочерних цепей, комплементарных и антипараллельных нитям материнской ДНК.
Механизма репликации – полуконсервативный!
3. Повреждения ДНК и их исправления (репарация)
Репарация – особая функция клеток, заключающаяся в способности исправлять химические повреждения и разрывы в молекулах ДНК, поврежденной при нормальном биосинтезе ДНК в клетке или в результате воздействия физических или химических агентов. Нарушения в структуре цепи ДНК быстро определяются специальными ферментами, пораженный участок удаляется экзонуклеазами, заполняется ДНК-полимеразой I (β) и сшивается ДНК-лигазой. Ряд наследственных болезней, к примеру, пигментная ксеродерма, связана с нарушениями системы репарации.

4.Биосинтез РНК на ДНК-матрице (транскипция
Транскрипциейназывается синтез РНК на ДНК-матрице. В результате образуются первичные траскрипты (пре-мРНК, пре-тРНК, пре-рРНК), комплементарные матричной цепи ДНК, имеющей направление от 3'- к 5'-концу. Субстратами и источниками энергии для синтеза РНК являются рибонуклеозидтрифосфаты (НТФ: АТФ, ГТФ, ЦТФ, УТФ).
Катализируют синтез РНК ферменты РНК-полимеразы. В ядре клеток эукариотов обнаружены три фермента:
• РНК-полимераза I, синтезирующая пре-рРНК;
• РНК-полимераза II, ответственная за синтез пре-мРНК;
• РНК-полимераза III, синтезирующая пре-тРНК.
На всех этапах транскрипции необходимо участие специальных белковых факторов: ТАТА-фактор, G, σ, А, В – факторов инициации, Е, Н, F – факторов элонгации, ρ-фактор – фактор терминации.
В основе процесса лежит принцип комплементарности оснований в полинуклеотидной цепи матричной ДНК и синтезируемой РНК, когда против А встает У, против Г - Ц, а против Т - А.
Суммарное уравнение синтеза РНК можно представить следующим образом:
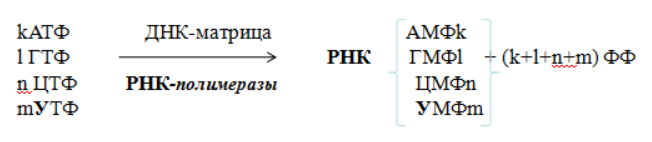
Биосинтез РНК происходит в участке ДНК, который называется транскриптон, с одного края он ограничен промотором (начало), с другого – терминатором (конец).
РНК-полимеразы эукариот имеют по две большие субъединицы и несколько малых субъединиц.
В процессе транскрипции различают стадии инициации, элонгации и терминации (рис. 3).
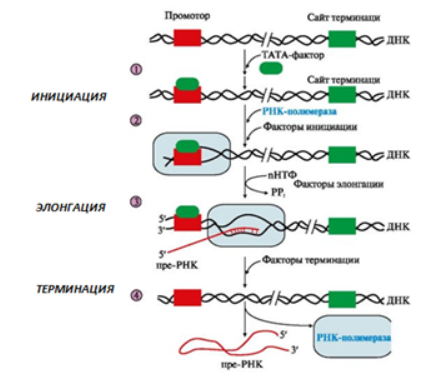
Рис. 3. Стадии транскрипции:
-инициация:
1)присоединение в область промотора белка, который называется «ТАТА-фактор»;
2) включение РНК-полимеразы в промоторный участок, при этом в зоне присоединения РНК-полимеразы происходит локальное расплетение двойной спирали ДНК;
-элонгация
:
3) рост нити пре-РНК;
-терминация:
4)освобождение в сайте терминации пре-РНК и РНК-полимеразы из комплекса с ДНК ускоряют факторы терминации.
Посттранскрипционные модификации РНК (процессинг)
Прежде чем выйти из ядра, каждый первичный транскрипт после ряда ковалентных модификаций превращается в «зрелую» молекулу РНК. К посттрансляционным модификациях относят реакции: кэпирования, полиаденилирования, сплайсинг, модификация азотистых оснований, метилирование и другие (рис. 4, 5).
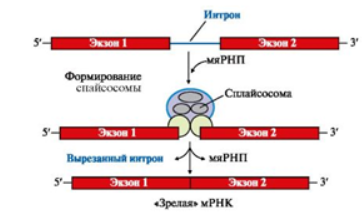
Рис. 4. Сплайсинг пре-мРНК
В ядре интроны удаляются при участии малых ядерных рибонуклеопротеинов (мяРНК), которые образуют комплекс, или сплайсосому. Плайсосомы гидролизуют 3´,5´-фосфодиэфирные связи на границе интрон-экзон и связывают экзоны между собой. Ферментативно активностью обладают РНК в составе мяРНП.
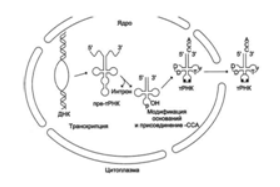
Рис. 5. Созревание пре-тРНК.
Первоначально в ядре: 1-удаляются участки полинуклеотидной цепи на 5´- и 3´-концах молекулы пре-тРНК, интрон в центральной области; 2-модифицируются азотистые основания и к 3´-концу присоединяется триплет ССА; 3-зрелые тРНК выходят из ядра в цитоплазму.
В синтезе РНК большую роль играет участок ДНК, который называется промотором. Какова его функция? Связывает РНК-полимеразу.
Связывает РНК-полимеразу. Какая схема соответствует главному постулату, или центральной догме, молекулярной биологии? Связывает РНК-полимеразуКакая схема соответствует главному постулату, или центральной догме, молекулярной биологии?ДНК→ мРНК → белок

