Файл: Контрольная работа по дисциплине Генетика Группа Шифр б проверил профессор Дюкова Н. Н. Тюмень, 2022.docx
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 03.05.2024
Просмотров: 47
Скачиваний: 1
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
Министерство сельского хозяйства Российской Федерации
ФГОУ ВО ГАУ Северного Зауралья
Институт дистанционного образования
Кафедра общей биологии
Контрольная работа по дисциплине Генетика
Выполнил:
Группа
Шифр Б-
Проверил: профессор Дюкова Н.Н.
Тюмень, 2022
ОГЛАВЛЕНИЕ
17. Генетический контроль синтеза ДНК. Репликация. Особенности репликации ДНК у эукариотов и прокариотов 3
44. Модификационная изменчивость, методы ее изучения. Длительные модификации и морфозы 10
63. Инбридинг, его генетическая сущность. Роль инбридинга в эволюции и селекции 12
71. У томата красная окраска плодов доминирует над желтой окраской. От самоопыления гетерозиготного по признаку окраски плодов томата получено потомство. Определите фенотип и генотип потомства 14
79. У томатов высокий рост доминирует над низким ростом, красная окраска плодов - над желтой окраской. Растения F1 были скрещены с чистосортными растениями, имеющими все признаки в доминантном состоянии. В F1 было получено 81 растение. Написать схему скрещивания и провести генетический анализ. 15
87. У собак доминантный аллель гена А обусловливает черную масть, рецессивный аллель, а - коричневую. Доминантный ген-ингибитор I подавляет проявление действия обоих генов и обусловливает белую масть. Рецессивный аллель гена-ингибитора i не оказывает влияния на окраску шерсти. При скрещивании гомозиготных собак белой и коричневой масти, имеющих генотипы ААII и ааii получили 24 щенка в F1 и 48 - в F2. Написать схему скрещивания и провести генетический анализ 16
98. При апробации табака установили частоту доминантного гена устойчивости к черной корневой гнили (р = 0.98). Определите фенотипическую и генотипическую структуру популяции табака 17
17. Генетический контроль синтеза ДНК. Репликация. Особенности репликации ДНК у эукариотов и прокариотов
Синтез ДНК – это естественное или искусственное создание молекул дезоксирибонуклеиновой кислоты (ДНК). ДНК – это макромолекула, состоящая из нуклеотидных звеньев, которые связаны ковалентными связями и водородными связями в повторяющейся структуре. Синтез ДНК происходит, когда эти нуклеотидные единицы соединяются с образованием ДНК; это может происходить искусственно (in vitro) или естественным путем (in vivoНуклеотидные единицы состоят из азотистого основания (цитозина, гуанина, аденина или тимина), пентозного сахара (дезоксирибозы) и фосфатной группы. Каждая единица соединяется, когда образуется ковалентная связь между ее фосфатной группой и пентозным сахаром следующего нуклеотида, образуя сахаро-фосфатный остов. ДНК является комплементарной, двухцепочечной структурой, поскольку специфическое спаривание оснований (аденин и тимин, гуанин и цитозин) происходит естественным образом, когда между нуклеотидными основаниями образуются водородные связи.
Существует несколько различных определений синтеза ДНК: это может относиться к репликации ДНК - биосинтезу ДНК (амплификация ДНК in vivo), полимеразной цепной реакции – ферментативному синтезу ДНК (амплификация ДНК in vitro) или синтезу генов - физическому созданию искусственных последовательностей генов. Хотя каждый тип синтеза очень отличается, у них есть некоторые общие черты. Нуклеотиды, которые были соединены с образованием полинуклеотидов может действовать как ДНК-шаблон для одной из форм синтеза ДНК - ПЦР. Репликация ДНК также работает с использованием шаблона ДНК, двойная спираль ДНК разматывается во время репликации, подвергая непарные основания для новых нуклеотидов водородной связи. Однако для синтеза генов не требуется матрица ДНК, и гены собираются de novo. Синтез ДНК происходит у всех эукариот и прокариот, а также у некоторых вирусов. Точный синтез ДНК важен для того, чтобы избежать мутаций ДНК. У людей мутации могут привести к таким заболеваниям, как рак, поэтому синтез ДНК и механизмы, задействованные in vivo, широко изучались на протяжении десятилетий. В будущем эти исследования могут быть использованы для разработки технологий, связанных с синтезом ДНК, которые будут использоваться в хранении данных.
Репликация (от лат. replicatio – возобновление) – процесс создания двух дочерних молекул ДНК на основе родительской молекулы ДНК. Репликацию ДНК осуществляет сложный комплекс, состоящий из 15-20 различных белков-ферментов, называемый реплисомой. С помощью специальных ферментов двойная спираль материнской ДНК расплетается на две нити, на каждой образовавшейся нити достраивается вторая нить, образуя две идентичных дочерних молекулы ДНК, которые затем скручиваются в отдельные спирали. В ходе последующего деления материнской клетки каждая дочерняя клетка получает по одной копии молекулы ДНК, которая является идентичной ДНК исходной материнской клетки. Этот процесс обеспечивает точную передачу генетической информации из поколения в поколение. РНК является самой важной частью этого процесса.
Репликация ДНК – ключевое событие в ходе деления клетки. Принципиально, чтобы к моменту деления ДНК была реплицирована полностью и при этом только один раз. Это обеспечивается определёнными механизмами регуляции репликации ДНК. Репликация проходит в три этапа:
-
инициация репликации -
элонгация -
терминация репликации.

Регуляция репликации осуществляется в основном на этапе инициации. Это достаточно легко осуществимо, потому что репликация может начинаться не с любого участка ДНК, а со строго определённого, называемого сайтом инициации репликации. В геноме таких сайтов может быть как всего один, так и много. С понятием сайта инициации репликации тесно связано понятие репликон. Репликон – это участок ДНК, который содержит сайт инициации репликации и реплицируется после начала синтеза ДНК с этого сайта. Геномы бактерий, как правило, представляют собой один репликон, это значит, что репликация всего генома является следствием всего одного акта инициации репликации. Геномы эукариот (а также их отдельные хромосомы) состоят из большого числа самостоятельных репликонов, это значительно сокращает суммарное время репликации отдельной хромосомы. Молекулярные механизмы, которые контролируют количество актов инициации репликации в каждом сайте за один цикл деления клетки, называются контролем копийности. В бактериальных клетках помимо хромосомной ДНК часто содержатся плазмиды, которые представляют собой отдельные репликоны. У плазмид существуют свои механизмы контроля копийности: они могут обеспечивать синтез как всего одной копии плазмиды за клеточный цикл, так и тысяч копий.
Репликация начинается в сайте инициации репликации с расплетания двойной спирали ДНК, при этом формируется репликационная вилка — место непосредственной репликации ДНК. В каждом сайте может формироваться одна или две репликационные вилки в зависимости от того, является ли репликация одно- или двунаправленной. Более распространена двунаправленная репликация. Через некоторое время после начала репликации в электронный микроскоп можно наблюдать репликационный глазок – участок хромосомы, где ДНК уже реплицирована, окружённый более протяжёнными участками нереплицированной ДНК. В репликационной вилке ДНК копирует крупный белковый комплекс (реплисома), ключевым ферментом которого является ДНК-полимераза. Репликационная вилка движется со скоростью порядка 100 000 пар нуклеотидов в минуту у прокариот и 500-5000 – у эукариот.
Ферменты (хеликаза, топоизомераза) и ДНК-связывающие белки расплетают ДНК, удерживают матрицу в разведённом состоянии и вращают молекулу ДНК. Правильность репликации обеспечивается точным соответствием комплементарных пар оснований и активностью ДНК-полимеразы, способной распознать и исправить ошибку. Репликация у прокариот осуществляется несколькими разными ДНК-полимеразами. ДНК-полимераза I действует на запаздывающей цепи для удаления РНК-праймеров и дорепликации очищенных мест ДНК. ДНК полимераза III – основной фермент репликации ДНК, осуществляющий синтез ведущей цепи ДНК и фрагментов Оказаки при синтезе запаздывающей цепи. Далее происходит закручивание синтезированных молекул по принципу суперспирализации и дальнейшей компактизации ДНК. Синтез энергозатратный.
Цепи молекулы ДНК расходятся, образуют репликационную вилку, и каждая из них становится матрицей, на которой синтезируется новая комплементарная цепь. В результате образуются две новые двуспиральные молекулы ДНК, идентичные родительской молекуле. Характеристики процесса репликации
-
матричный – последовательность синтезируемой цепи ДНК однозначно определяется последовательностью материнской цепи в соответствии с принципом комплементарности; -
полуконсервативный – одна цепь молекулы ДНК, образовавшейся в результате репликации, является вновь синтезированной, а вторая – материнской; -
идёт в направлении от 5’-конца новой молекулы к 3’-концу; -
полунепрерывный – одна из цепей ДНК синтезируется непрерывно, а вторая – в виде набора отдельных коротких фрагментов (фрагментов Оказаки); -
начинается с определённых участков ДНК, которые называются сайтами инициации репликации (англ. origin).
Благодаря своей 3'-5' – экзонуклеазной гидролитической активности ДНК-полимераза может исключить неправильный нуклеотид из цепочки и затем вставить на его место правильный, после чего репликация продолжается в нормальном режиме. У прокариот имеются специальные терминаторы, прекращающие синтез цепи ДНК. Участок ДНК у E. coli, на котором происходит терминация, имеет длину 350 тысяч пар нуклеотидов (350 килобаз). После удаления праймеров и застраивания брешей ДНК-полимеразой отдельные участки дочерней цепи ДНК сшиваются между собой ферментом ДНК-лигазой.Непрерывная сборка идет быстрее, чем фрагментарная. Поэтому одна из дочерних цепей ДНК называется лидирующей, или ведущей, вторая – запаздывающей, или отстающей. У прокариот репликация протекает быстрее: примерно 1000 нуклеотидов в секунду. В то время как у эукариот только около 100 нуклеотидов. Количество нуклеотидов в каждом фрагменте Оказаки у эукариот составляет примерно до 200, у прокариот – до 2000. У прокариот кольцевые молекулы ДНК представляют собой один репликон. У эукариот каждая хромосома может содержать множество репликонов. Поэтому синтез начинается в нескольких точках, одновременно или нет. Ферменты и другие белки репликации действуют совместно, образуя комплекс и двигаясь по ДНК. Всего в процессе участвует около 20 разных белков, здесь были перечислены лишь основные.
В отличие от прокариот, имеющих пять ДНК-полимераз, эукариоты содержат по меньшей мере пятнадцать ДНК-полимераз, которые обозначаются буквами греческого алфавита. Основными ДНК-иолимеразами считаются шесть ферментов, три из которых (альфа – а, дельта – б и эпсилон – е) непосредственно участвуют в репликации хромосомной ДНК. ДНК-полимсраза бета (р) обеспечивает репарацию ДНК, вырезая поврежденные основания и застраивая возникшие бреши. ДНК-полимераза гамма (у) реплицирует и обеспечивает репарацию митохондриальной ДНК, имея 3' –» 5' экзонуклеазную активность. ДНК-полимераза дзета (Q способна использовать в качестве матрицы ДНК, содержащую циклобутановые димеры. ДНК-полимераза эта (г|), так же, как и ДНК-полимераза дзета, вовлекается в 505-ответ клетки на ДНК-повреждающее воздействие. Теломераза – специфическая обратная транскриптаза, катализирующая синтез теломерных концов хромосом. Целый ряд эукариотических ДНК- полимераз (0, А, ф, ст и р) пока слабо охарактеризован.
Эукариотическая ДНК-праймаза, в отличие от аналогичного белка прокариот, образует постоянный комплекс с ДНК-полимеразой альфа, роль которого, очевидно, ограничивается синтезом праймеров при репликации обеих цепей ДНК. Механизмы репликации ДНК прокариот и эукариот существенно различаются в том отношении, что во втором случае синтез ведущей и отстающей цепей ДНК осуществляют разные ДНК-полимеразы (епсилон и дельта соответственно), тогда как у прокариот обе цепи ДНК синтезируются димером ДНК-полимеразы III.
В отличие от бактерий, у которых процесс репликации начинается лишь в одной-двух точках, в эукариотической клетке инициация репликации происходит на специфических множественных последовательностях нуклеотидов – репликаторах, которые расположены на расстоянии около 100000 пн. По хромосоме эукариот в каждый данный момент может двигаться независимо друг от друга множество репликативных вилок, что и позволяет в короткий срок реплицировать всю хромосому. Частота использования отдельных репликаторов изменяется в онтогенезе, уменьшаясь в клетках взрослого организма. Интересно, что на этапе инициации репликации эукариот происходит взаимодействие регуляторных последовательностей репликатора по меньшей мере с шестью различными белками, которые образуют комплекс, распознающий область начала репликации.
Удвоение ДНК у эукариот проходит в S-фазу клеточного цикла. Инициацию репликации регулируют специфические сигнальные белковые молекулы – факторы роста. Они связываются с рецепторами клеточных мембран, генерируя сигнал, который и побуждает клетку к началу репликации. Одними из первых активируются гены, кодирующие белки циклины. Циклинзависимые киназы, связывая циклин, переходят в активную форму и фосфорилируют специфические белки, которые регулируют синтез ферментов, обеспечивающих репликацию.
Синтез новых цепей ДНК может произойти только при расхождении родительских цепей. В точке начала репликаци (сайты инициации или ориджины) происходит локальное расхождение цепей ДНК и образуются две репликативные вилки, движущиеся в противоположных направлениях.
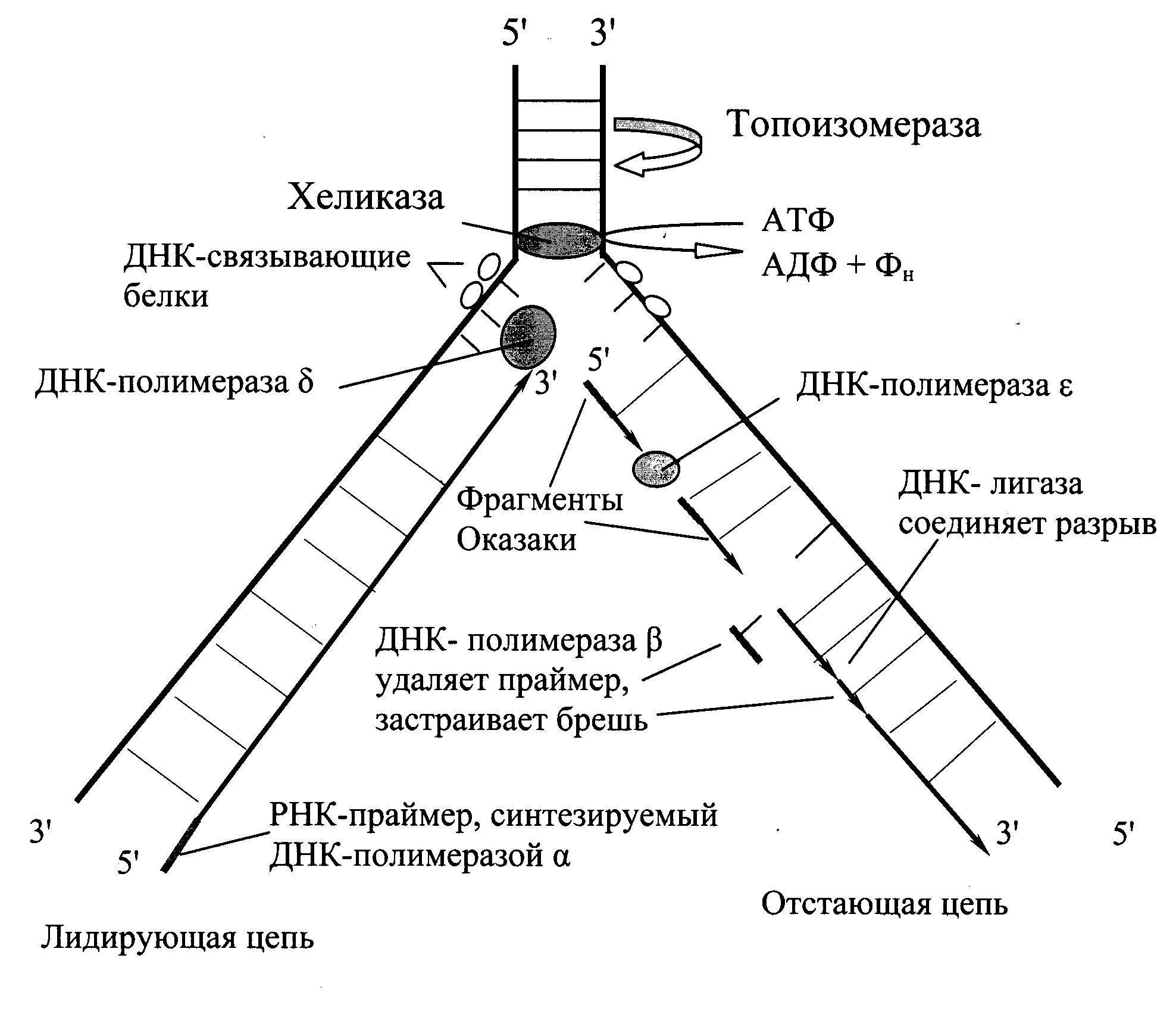
Рис.1. Репликация ДНК.
После окончания репликации происходит метилирование вновь образованных цепей ДНК. Наличие СН3-групп необходимо для формирования структуры хромосом, а также для регуляции транскрипции генов.