Файл: Федеральное государственное бюджетное образовательное учреждение высшего образования астраханский государственный университет.docx
Добавлен: 03.05.2024
Просмотров: 34
Скачиваний: 0
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
СОДЕРЖАНИЕ
1.1 Общая характеристика фотосинтеза
Глава 2 ФОТОСИНТЕТИЧЕСКИЕ ПИГМЕНТЫ
2.1 Структурная основа для фотосинтетических процессов
2.3 Спектры поглощения хлорофиллов
3.1 Физико-химическая основа фотосинтеза.
3.2 БИОФИЗИЧЕСКИЕ МЕХАНИЗМЫ ФОТОСИНТЕЗА.
3.3. Основные компоненты электротранспортной цепи
3.4. Циклический и псевдоциклический транспорт электрона
МИНИСТЕРСТВО ОБРАЗОВАНИЯ И НАУКИ РОССИЙСКОЙ ФЕДЕРАЦИИ
ФЕДЕРАЛЬНОЕ ГОСУДАРСТВЕННОЕ БЮДЖЕТНОЕ
ОБРАЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ ВЫСШЕГО ОБРАЗОВАНИЯ
«АСТРАХАНСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ»
Биологический факультет
Кафедра физиологии, морфологии, генетики и биомедицины
Курсовая работа выполнена в рамках изучения дисциплины «Биофизика»
«Биофизика фотосинтеза»
Направление подготовки: 06.03.01 Биология
Направленность (профиль): Генетика
Выполнила:
Мухтарова Амина Рауфовна
Научный руководитель:
кандидат биологических наук;
заместитель декана биологического факультета;
доцент кафедры
Касимова Сауле Куаншевна
Астрахань-2019
Оглавление
МИНИСТЕРСТВО ОБРАЗОВАНИЯ И НАУКИ РОССИЙСКОЙ ФЕДЕРАЦИИ 1
3
Глава 1 ОБЩИЕ ПОНЯТИЯ 4
1.1 Общая характеристика фотосинтеза 4
1.2 Изучение фотосинтеза 4
1.3 Значение фотосинтеза 5
Глава 2 ФОТОСИНТЕТИЧЕСКИЕ ПИГМЕНТЫ 6
2.1 Структурная основа для фотосинтетических процессов 6
2.2 Светособирающие комплексы 7
2.2.1 Фотосинтетические пигменты 8
2.3 Спектры поглощения хлорофиллов 9
Глава 3 ПРОЦЕССЫ ФОТОСИНТЕЗА 10
3.1 Физико-химическая основа фотосинтеза. 10
3.2 БИОФИЗИЧЕСКИЕ МЕХАНИЗМЫ ФОТОСИНТЕЗА. 15
3.3. Основные компоненты электротранспортной цепи 16
3.3.1. Фотосистема I 16
3.3.2. Фотосистема II 16
3.3.3. b6f или b/f-комплекс 18
3.4. Циклический и псевдоциклический транспорт электрона 18
3.5 Основные свойства фотосинтетического аппарата растений 18
3.5.1 Насыщение световой кривой фотосинтеза с ростом освещенности 18
3.5.2 «Эффект усиления» Эммерсона. 19
3.5.3 Эффект Кока-Жолио. 20
3.5.4 Задержанная флуоресценция. 21
ЗАКЛЮЧЕНИЕ 22
Литература 23
ВВЕДЕНИЕ
Фотосинтез - один из важнейших механизмов жизни на Земле, поскольку, за его счет образуются как сложные высокоупорядоченные органические вещества, так и кислород, потребляемые другими организмами (бактериями, растениями и животными) в качестве источников строительного материала и энергии.
В работе мы более подробно рассматриваем механизм фотосинтеза, а именно обращаем внимание происходящие процессы со стороны биофизики. Фотосинтез – сложный, но вместе с тем интересный и очень красивый процесс. Он появился на ранних этапах эволюции у различных бактерий – пурпурных, цианобактерий, зеленых серных бактерий и других, у одноклеточных эукариот (водорослей) и, наконец, у высших растений. Исследование фотосинтетического аппарата прокариот внесло большой вклад в выяснение основных принципов и механизмов фотосинтеза. Но здесь мы рассмотрим главным образом механизмы фотосинтеза высших растений.
Суть фотосинтеза заключается в получении высокоэнергетических электронов за счет энергии квантов света, поглощаемых хлорофиллом. Для того, чтобы предупредить возвращение электрона в исходное состояние и растрату полученной энергии на тепловые процессы, возбужденный электрон очень быстро переносится по электронтранспортной цепи на NADP+, который затем используется в биосинтетических реакциях. Исходным источником электронов является вода, которая при этом расщепляется и выделяет кислород. Этим кислородом, являющимся побочным продуктом фотосинтеза, дышат все аэробные организмы.
Глава 1 ОБЩИЕ ПОНЯТИЯ
1.1 Общая характеристика фотосинтеза
Фотосинтез (от греч. φωτο- — свет и σύνθεσις — синтез, совмещение, помещение вместе) — процесс образования органических веществ из углекислого газа и воды на свету при участии фотосинтетических пигментов (хлорофилл у растений, бактериохлорофилл и бактериородопсин у бактерий). В современной физиологии растений под фотосинтезом чаще понимается фотоавтотрофная функция — совокупность процессов поглощения, превращения и использования энергии квантов света в различных эндэргонических реакциях, в том числе превращения углекислого газа в органические вещества.
1.2 Изучение фотосинтеза
Первые опыты по фотосинтезу были проведены Джозефом Пристли в 1770—1780-х годах, когда он обратил внимание на «порчу» воздуха в герметичном сосуде горящей свечой (воздух переставал быть способен поддерживать горение, помещённые в него животные задыхались) и «исправление» его растениями. Пристли сделал вывод что растения выделяют кислород, который необходим для дыхания и горения, однако не заметил что для этого растениям нужен свет. Это показал вскоре Ян Ингенхауз. Позже было установлено, что помимо выделения кислорода растения поглощают углекислый газ и при участии воды синтезируют на свету органическое вещество. В 1842 Роберт Майер на основании закона сохранения энергии постулировал, что растения преобразуют энергию солнечного света в энергию химических связей. В 1877 В. Пфеффер назвал этот процесс фотосинтезом. Хлорофиллы были впервые выделены в 1818 П. Ж. Пельтье и Ж. Кавенту. Разделить пигменты и изучить их по отдельности удалось М. С. Цвету с помощью созданного им метода хроматографии. Спектры поглощения хлорофилла были изучены К. А. Тимирязевым, он же, развивая положения Майера, показал что именно поглощенные позволяют повысить энергию системы, создав вместо слабых связей С-О и О-Н высокоэнергетические С-С (до этого считалось что в фотосинтезе используются жёлтые лучи, не поглощаемые пигментами листа). Сделано это было благодаря созданному им методу учёта фотосинтеза по поглощённому CO2, в ходе экспериментов по освещению растения светом разных длин волн (разного цвета) оказалось что интенсивность фотосинтеза совпадает со спектром поглощения хлорофилла. Окислительно-восстановительную сущность фотосинтеза (как оксигенного, так и аноксигенного) постулировал Корнелис ван Ниль. Это означало, что кислород в фотосинтезе образуется полностью из воды, что экспериментально подтвердил в 1941 А. П. Виноградов в опытах с изотопной меткой. В 1937 г. Роберт Хилл установил, что процесс окисления воды (и выделения кислорода), а также ассимиляции CO2 можно разобщить. В 1954—1958 Д. Арнон установил механизм световых стадий фотосинтеза, а сущность процесса ассимиляции CO2 была раскрыта Мельвином Кальвином с использованием изотопов углерода в конце 1940-х, за эту работу в 1961 ему была присуждена Нобелевская премия. В 1955 году был выделен и очищен фермент рибулозобисфосфат-карбоксилаза/оксигеназа.С4 фотосинтез был описан Ю. С. Карпиловым в 1960 и М. Д. Хэтчем и К. Р. Слэком в 1966.

1.3 Значение фотосинтеза
Фотосинтез является основным источником биологической энергии, фотосинтезирующие автотрофы используют её для синтеза органических веществ из неорганических, гетеротрофы существуют за счёт энергии, запасённой автотрофами в виде химических связей, высвобождая её в процессах дыхания и брожения. Энергия, получаемая человечеством при сжигании ископаемого топлива (уголь, нефть, природный газ, торф), также является запасённой в процессе фотосинтеза.
Фотосинтез является главным входом неорганического углерода в биологический цикл. Весь свободный кислород атмосферы — биогенного происхождения и является побочным продуктом фотосинтеза. Формирование окислительной атмосферы (кислородная катастрофа) полностью изменило состояние земной поверхности, сделало возможным появление дыхания, а в дальнейшем, после образования озонового слоя, позволило жизни выйти на сушу.
Глава 2 ФОТОСИНТЕТИЧЕСКИЕ ПИГМЕНТЫ
2.1 Структурная основа для фотосинтетических процессов
Фотосинтетические процессы в листьях локализованы в специальных органеллах, называемых хлоропластами. Внутреннее пространство хлоропласта заполнено пластинчатыми гранулами зеленого цвета – гранами. Граны представляют собой стопки сильно сплюснутых мешочков – тилакоидами. Стенки тилакоидов – ламеллы – представляют собой тонкие фосфолипидные мембраны, внутри которых находятся липотропные участники первичных процессов фотосинтеза. К ним относятся белки, пигменты, переносчики электронов и протонов. Внутреннее межмембранное пространство тилакоидов является водной средой, в которой содержатся водорастворимые участники процесса. В межгранальном – стромальном – пространстве (также водном), находятся менее упакованные отдельные тилакоиды и ламеллы.(Рис.1)
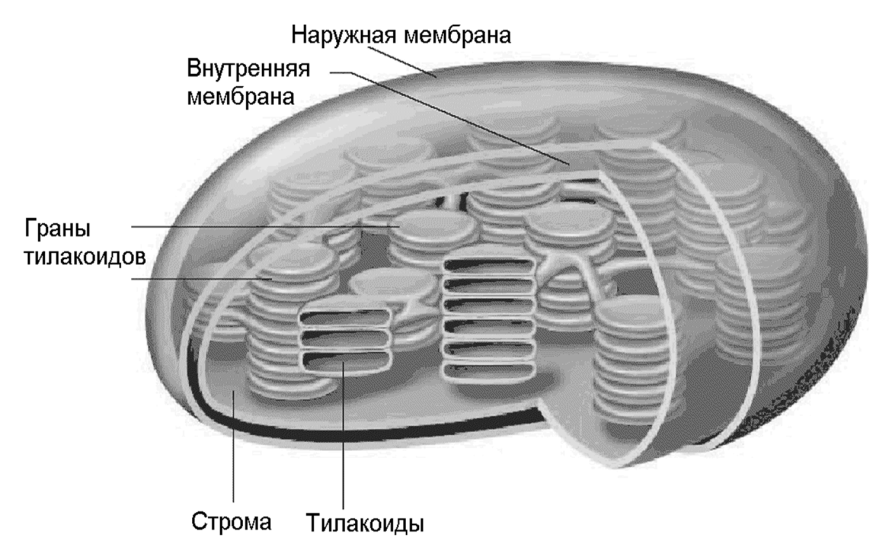
Рис.1. Строение хлоропласта
2.2 Светособирающие комплексы
Хлорофилл выполняет две функции: поглощения и передачи энергии. Более 90 % всего хлорофилла хлоропластов входит в состав светособирающих комплексов (ССК), выполняющих роль антенны, передающей энергию к реакционному центру фотосистем I или II. Помимо хлорофилла в ССК имеются каротиноиды, а у некоторых водорослей и цианобактерий — фикобилины, роль которых заключается в поглощении света тех длин волн, которые хлорофилл поглощает сравнительно слабо.Передача энергии идёт резонансным путём (механизм Фёрстера) и занимает для одной пары молекул 10−10—10−12 с, расстояние на которое осуществляется перенос составляет около 1 нм. Передача сопровождается некоторыми потерями энергии (10 % от хлорофилла a к хлорофиллу b, 60 % от каратиноидов к хлорофиллу), из-за чего возможна только от пигмента с максимумом поглощения при меньшей длине волны к пигменту с большей. Именно в таком порядке взаимно локализуются пигменты ССК, причём наиболее длинноволновые хлорофиллы находятся в реакционных центрах. Обратный переход энергии невозможен.ССК растений расположен в мембранах тилакоидов, у цианобактерий основная его часть вынесена за пределы мембран в прикреплённые к ним фикобилисомы — палочковидные полипептидно-пигментные комплексы, в которых находятся различные фикобилины: на периферии фикоэритрины (с максимумом поглощения при 495—565 нм), за ними фикоцианины (550—615 нм) и аллофикоцианины (610—670 нм), последовательно передающие энергию на хлорофилл a (680—700 нм) реакционного центра.
2.2.1 Фотосинтетические пигменты
Основной пигмент фотосинтеза - хлорофилл а (Chl a). Он состоит из порфириновой головки размером примерно 1,5х1,5 нм, обладающей гидрофильными свойствами, и длинного гидрофобного хвоста - фитола - длиной около 2,0 нм (Рис.2). В центре порфиринового кольца хлорофиллов находится ион Mg2+, связанный координационными связями с атомами азота (Рис.2). Другие формы хлорофилла - b,c,d, а также бактериохлорофилл - отличаются боковыми заместителями и распределением двойных связей в порфириновом кольце.

Рис.2. Структура хлорофилла а. Отмечены модификации, характерные для хлорофилла b и бактериохлорофилла.
2.3 Спектры поглощения хлорофиллов
Спектры поглощения хлорофиллов имеют две интенсивные полосы: характерную для порфиринов полосу Сорэ в синей или ближней ультрафиолетовой части спектра и «красную» полосу поглощения света в области 650-700 нм (у бактериохлорофиллов она смещена в ближнюю инфракрасную область 800-890 нм). Поглощение света в красной области спектра обеспечивается взаимодействием иона магния с сопряженной *-электронной системой порфиринового кольца. (Для сравнения - гем, содержащий в порфириновом кольце не Mg2+, а Fe2+ поглощает свет не в красной, а зелено-желтой части спектра). В клетке "красные полосы" поглощения хлорофиллов сдвинуты в длиноволновую область по сравнению со спектрами, полученными в органических растворителях (Табл.1), что указывает на значительную роль пигмент-пигментных, пигмент-белковых и пигмент-липидных взаимодействий вследствие плотной упаковки хлорофилльных молекул в фотосинтетической мембране.Методами низкотемпературной и дифференциальной спектрофотометрии выявлено до 10 модификаций хлорофилла a, различающихся по спектрам поглощения и флуоресценции (т.е. по расположению энергетических уровней), по экстрагируемости из мембраны (т.е. по гидрофильности/гидрофобности) и т.д. Они называются "нативными формами" хлорофилла а. Так, выделены Р670 (поглощающий свет 670-673 нм), Р680, Р700 и т.д. Основным фактором, обуславливающим свойства этих нативных форм, являются пигмент-пигментные взаимодействия, т.е. образование димеров и более крупных агрегатов. В пигментных агрегатах за счет взаимодействия *-электронных систем происходит "размазывание" электронного облака по двум или нескольким порфириновым кольцам. Соответственно, снижается верхний энергетический уровень и повышается способность поглощать более длинноволновый красный свет.
Таблица 1.
Полосы поглощения некоторых хлорофиллов в растворах органических растворителей и в клетках.
| Хлорофиллы | Органические растворители | Клетка | ||||
| Полоса Сорэ | Красная полоса | Полоса Сорэ | Красная полоса | | ||
| Хлорофилл а | 420 | 660 | 435 | 670-700 | | |
| Хлорофилл b | 450 | 640 | 480 | 650 | | |
| Хлорофилл c | 445 | 625 | | 645 | | |
| Хлорофилл d | 450 | 690 | | 740 | | |
| Бактериохлорофилл а | 365 | 605, 770 | - | 800-890 | | |
Кроме хлорофилла в фотосинтезирующих клетках встречаются и другие пигменты: феофитины, каротиноиды, фикоэритрины фикобилины и др. Все они содержат длинные цепочки сопряженных связей, в которых π-электронное облако «размазывается» по всей длине, что также приводит к снижению возбужденных энергетических уровней и позволяет поглощать кванты видимого света. У каротиноидов длина цепочек составляет примерно 40 углеродных атомов. Они поглощают свет в области 400-500 нм и имеют желтый или оранжевый цвет. Фикобилины поглощают свет в области 500-650 нм. Все они дополняют спектр поглощения хлорофилла, поглощая не только красный свет, но и свет других участков спектра, позволяя растениям эффективно улавливать солнечную радиацию.
Глава 3 ПРОЦЕССЫ ФОТОСИНТЕЗА
3.1 Физико-химическая основа фотосинтеза.
Основная продукция фотосинтеза - это соединения с высоким восстановительным потенциалом – углеводы. Рабочим телом фотосинтетической машины в высших растениях и некоторых водорослях является вода. Процесс фотосинтеза включает в себя две стадии:
1. Фотолиз, физическая сущность которого заключается в разложении воды с помощью квантов света на кислород и водород, причем кислород является в этом процессе побочным продуктом;