ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 18.03.2024
Просмотров: 10
Скачиваний: 0
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.

ЖҮЙКЕ БИОХИМИЯСЫ
ОЖЖ БИОЭНЕРГЕТИКАСЫ
Қалыпты жағдайдағы энергия көздері – аэробты гликолиз. Глюкозаның транспортерлары ГЛЮТ-1 – астроциттер және олигодентроциттерге, ГЛЮТ-3 – нейрондарға, ГЛЮТ-5 – микроглия жасушаларына.
Патология жағдайында – кетонды денелер.
Мидың арнайы қызметтері негізін құрайтын энергия тәуелді негізгі процестер
| Қызметтері | Биохимиялық реакциялар | Энергия шығымы% |
| Иондық асиммитрияны қайта қалыптастыруға байланысты жүйке импульстарының өткізілуі | К+ және Nа+ - АТФ-азалық реакциялар | 40 |
| Синаптикалық құрылымдардың қызметі | Синаптикалық құрылымдардың липо- және гликопротеиндік кешендерінің, арнайы протеиндердің синтезі, медиаторлардың синтезі және метаболизмі, медиаторлардың сақталуы, тасымалы, бөлініп шығуы, кері қармалуы; рецепторлардың қызметі | 30-35 |
| Ақпараттардың сақталуы және қайта өңделуі (нейрологиялық жады) | Арнайы протеиндердің, нейропептидтердің, нуклеин қышқылдарының, липо- және гликопротеиндік кешендерінің синтезі | 20 шамасында |
| Аксоналдық және ретроградтық ағым | Нейромиозиндер, динеиндер, кинезиндер тобындағы жиырылғыш протеиндер қызметі кезіндегі АТФ гидролизі | 2-3 |
| Нейрондардың құрылымдық бірліктерінің белгілі кеңістіктегі бағытын қалыптастыру(аксон, дендрит) | Жиырылғыш және құрылымдық протеиндердің фосфорлануы | 1-2 |
Ескерту: Жекелеген қызметтердің энергия шығыны ми үшін арнайы барлық қызметтерге жұмсалатын жалпы шығынның проценттік үлесімен белгіленеді.
Дамып өсуші мида энергия көзі ретінде глюкоза, май қышқылдары , аминқышқылдары қолданылады
Мида,әсіресе дамып өсуші мида пентозофосфаттық цикл қарқынды жүреді.
Мидағы қанайналымының жергілікті бұзылысы кезіндегі энергиятапшылығы тығырынан шығу үшін астроциттерде гликоген синтезі жүреді.
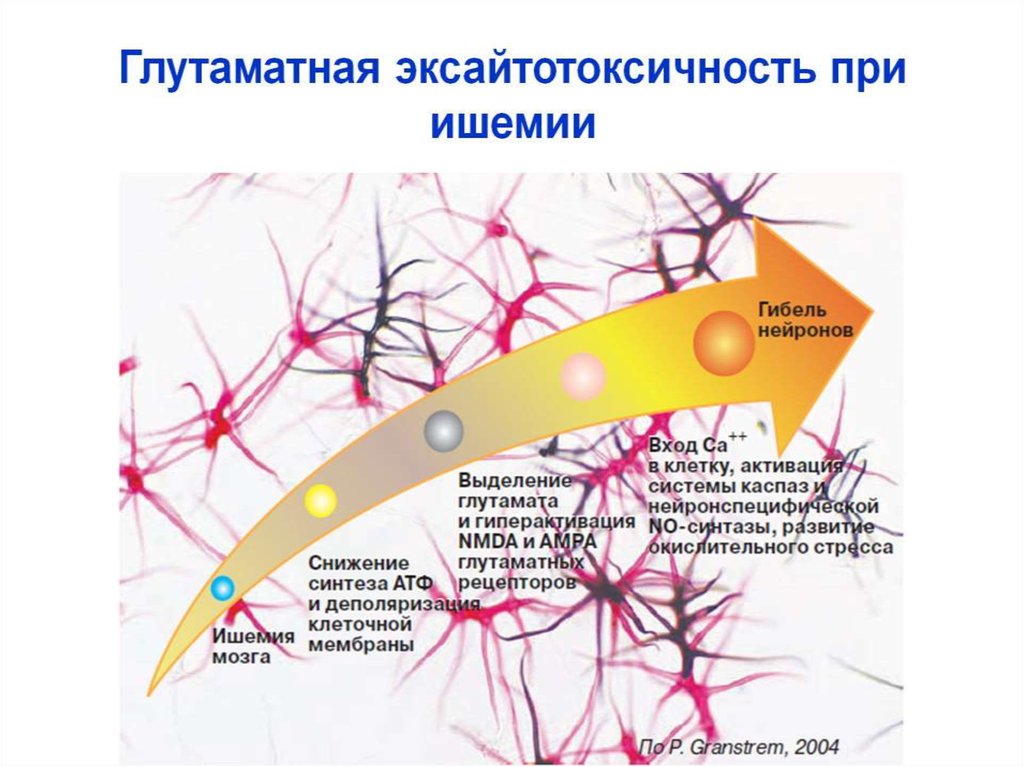
Жүйке ұлпасы гипоксияға өте сезімтал келеді..
Нейрондағы ҮКҚЦ маңызы: 1 энергетикалық; 2. Глутамат және ГАМК синтезі; 3. Цитрат цитоплазмаға тасымалданады – бос ацетил-КоА ацетилхолиннің синтезі үшін пайдаланылады.
ОЖЖ қалыпты жұмысы үшін қажетті жоғары қарқынды энергия алмасуын қамтамасыз ететін оттектің үнемі жеткізілуі қажетті жағдай болып табылады. Бас миы оттек тапшылығына өте сезімтал болады. Тыныс алу кезіндегі ауадағы оттектің парциалды қысымының 90 мм Нg (норма) - 25-30 мм Нg дейінгі төмендеуі коматоздық жағдайдың дамуына алып келеді. Миға оттектің жеткізілуінің күрт төмендеуі глюкозаның сіңірілуінің төмендеуімен қосарланады, және керісінше қанайналымындағы глюкоза деңгейінің кризистік мөлшерге (1,5-1,3 ммоль/л) дейін кемуі оттектің қолданылуын тежейді және кома дамуына (мысалы, инсулиндік кома)алып келеді.
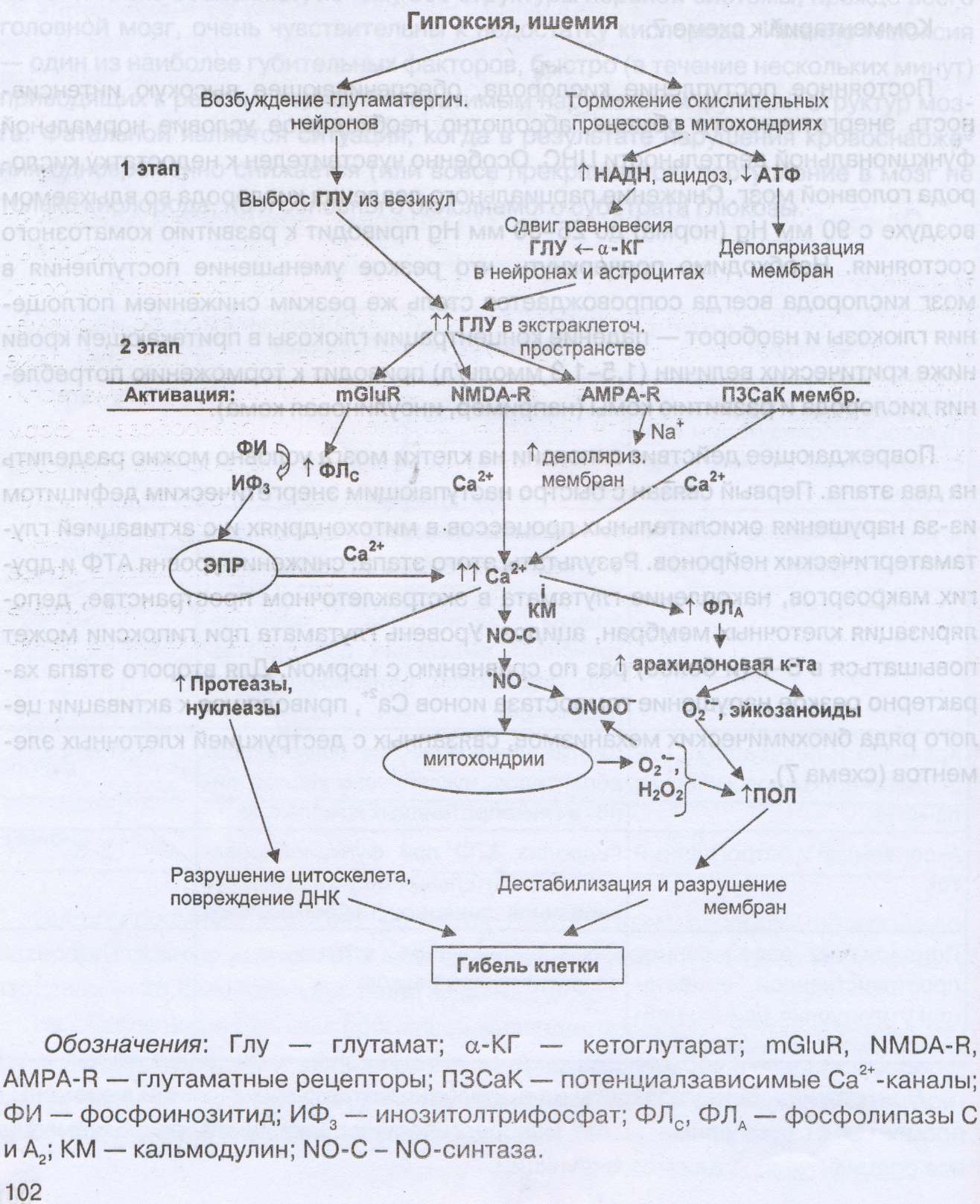
Ми жасушасына гипоксияның зақымдаушы әсері шартты түрде екі кезеңге бөлінеді.Біріншісі митохондрияларда тотығу процестерінің бұзылысынан шұғыл энерия тапшылығының пайда болуымен және глутаматергиялық нейрондардың белсендірілуімен байланысты болады. Бұл кезеңнің нәтижелері: АТФ және басқа да макроэртар деңгейінің төмендеуі, экстражасушалық кеңістікте глутаматтың жиналуы, жасуша мембранасының деполяризациясы, ацидоз. Гипоксия кезінде глутаматтың мөлшері қалыптымен салыстырғанда 5-7 (және одан артық ) кемуі мүмкін. Екінші кезеңі жасушалық элементтер деструкциясымен байланысты бірқатар биохимиялық механизмдердің белсендірілуіне алып келетін Са2+ иондарының гомеостазының күрт бұзылуымен сипатталады.
| Шығу көзі/Аминқышқылы | Нейромедиатор | Рецепторлар, синтезделу орны, синтездің ерекшеліктері | Роль | Тапшылық симптомдары | Артық мөлшерінің симптомдары | Агонист | Антагонист |
| Тирозин | Дофамин норадреналин | 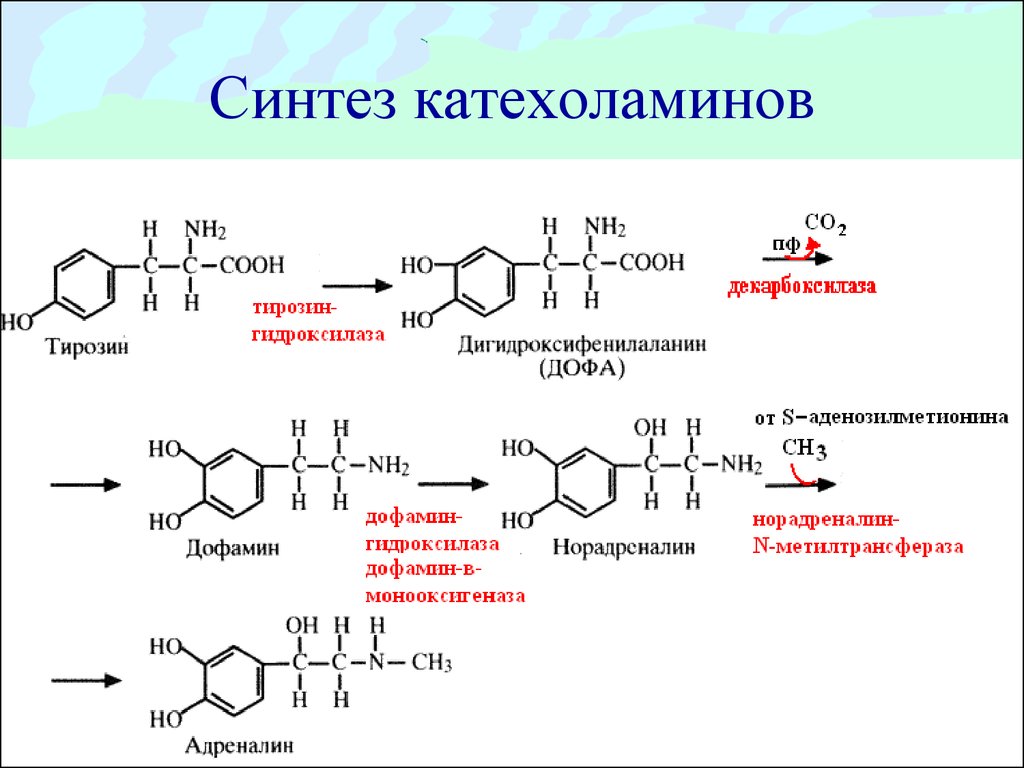 Рецепторы: D1, D2, D3, D4 – рецепторы дофамина Рецепторы – α1-, α2-, β1-, β2-адренорецепторы | Мотивация реакциялары, эмоционалдық реакциялар, рахаттану сезімі, мадақтаулар, қимыл-қозғалыс белсенділігі – ұсақ моторика, норадреналин – мидың жалпы белсендірілуі, оқудың жақсаруы, ауырсыну сезімінің төмендеуі, күйзеліс кезіндегі оң эмоциялар. | Паркинсонизм, депрессии | Ойлау қабілетінің бұзылысы, галлюцинация, шизофрения, қудалау маниясы | Кокаин Амфетамин, апоморфин, бромкриптин, | Хлорпромазин (аминазин), шалоперидол, |
| Глутамат | Глутамат | NMDA-рецепторлары | ОЖЖ негізгі қоздырушы медиаторы , жады | Импульстік мінез-құлық, аутизм | Эпилепсия, ми ишемиясы | Каинат | Кетамин |
| ГАМК | 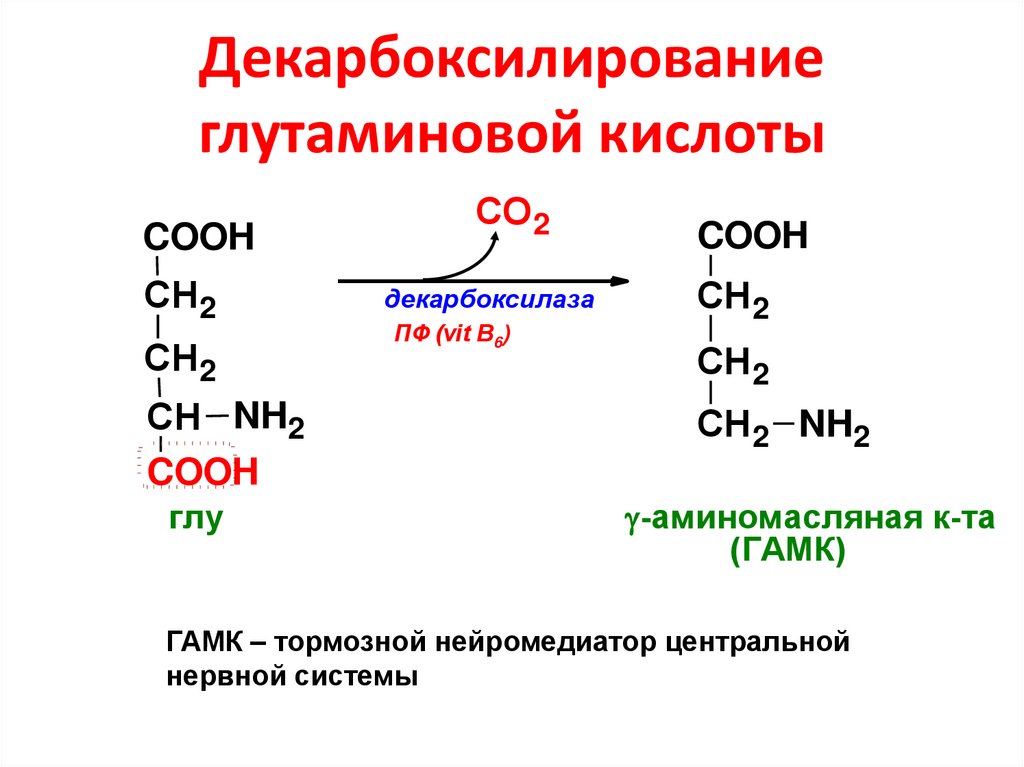 | Тежеуші нейромедиатор, ұйқы, жады | Эпилепсия, ұйқының бұзылуы, үрей | мазасыздану, ентікпе, қолдың қалтырауы | Барбитурат, бенздиазепины, баклофен | Бикукулин, пенициллин | |
| Триптофан | Серотонин | 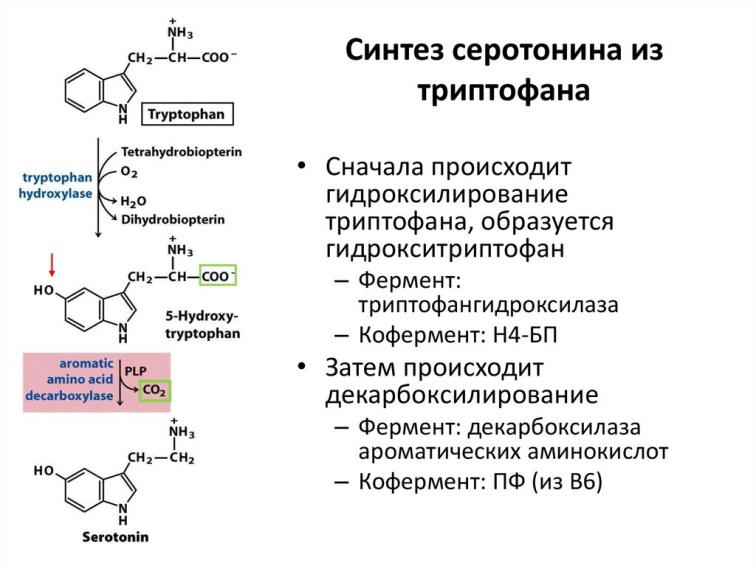 5НТ-1, 5НТ-2, 5НТ-3, 5НТ-4 | Терморегуляция, ұйқы, циркадтық ритм, депрессия, үрей, агрессия, жадының, көңіл- күйдің, тәбеттің реттелуі | депрессия | Эйфория, бас миы тамырларының тарылуы, мигрень | ЛСД | Хлорпромазин |
| Таурин | Таурин | | Балалардың жүйке жүйесінің дамуын қамтамасыз етеді, ұстама белсенділігін төмендетеді,жүйке жүйесінің қозуын және тежелуін реттейді | Эпилепсия | | шлицин | стрихнин |
| Серин Холин | Ацетилхолин | 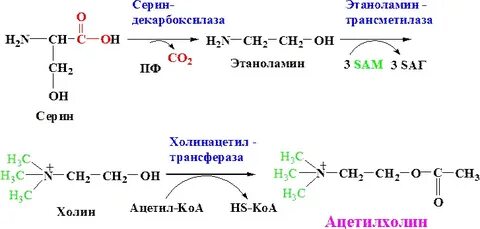 Н-/М-холинорецепторы | Жүйке- бұлшықет хабар таралуы, ұзақ қа созылған жады, ұйқы және сергектіктің реттелуі | Миастения | Судороги | Никотин | Атропин |
| Пептиды | Эндорфины энкефалины |  | | | | Морфин | Налоксон |
| | Анадамид | 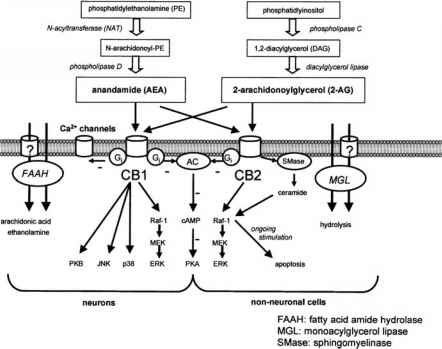 СВ1-, СВ2-рецепторы 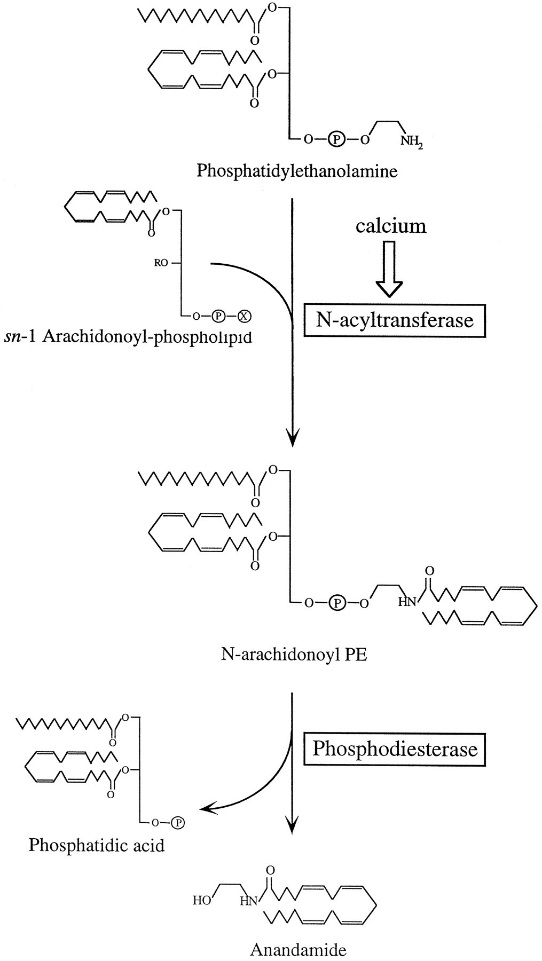 | Тәбеттің реттелуі, рахаттану сезімі, ауырсынудың басылуы, антидепрессант, тамаққа және темекіге тәуелділік, семіздік, Ауырсынудың, қорқыныштың, үрейдің сейілуі, зақымдалған нейрондардың жойылуынан сақтау, жүрек айнуы және құсуды бәсеңдету, тәбетті арттыру (т.н. «гедонистическая еда») | | | | Римонабант (Акомплиа) |
| АТФ | Аденозин | | Нейропротекция гипоксия және ми ишемиясы кезінде, глутамат рецепторларын басады, седатация, ұстамаға қарсы әрекет, жады | | | Аденозин | Кофеин, тнофиллин, |
 МИЕЛИННІҢ СИНТЕЗІ
МИЕЛИННІҢ СИНТЕЗІМИЕЛИННІҢ МОЛЕКУЛАЛЫҚ ҰЙЫМДАСУЫ
Миелиннің құрылымы туралы негізгі ақпарат рентген-құрылымдық анализ және электрондық микроскопия көмегімен алынады..
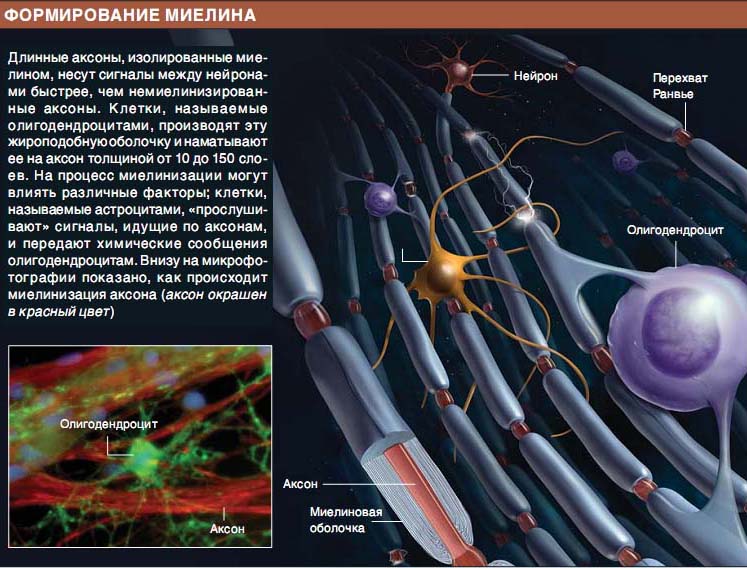
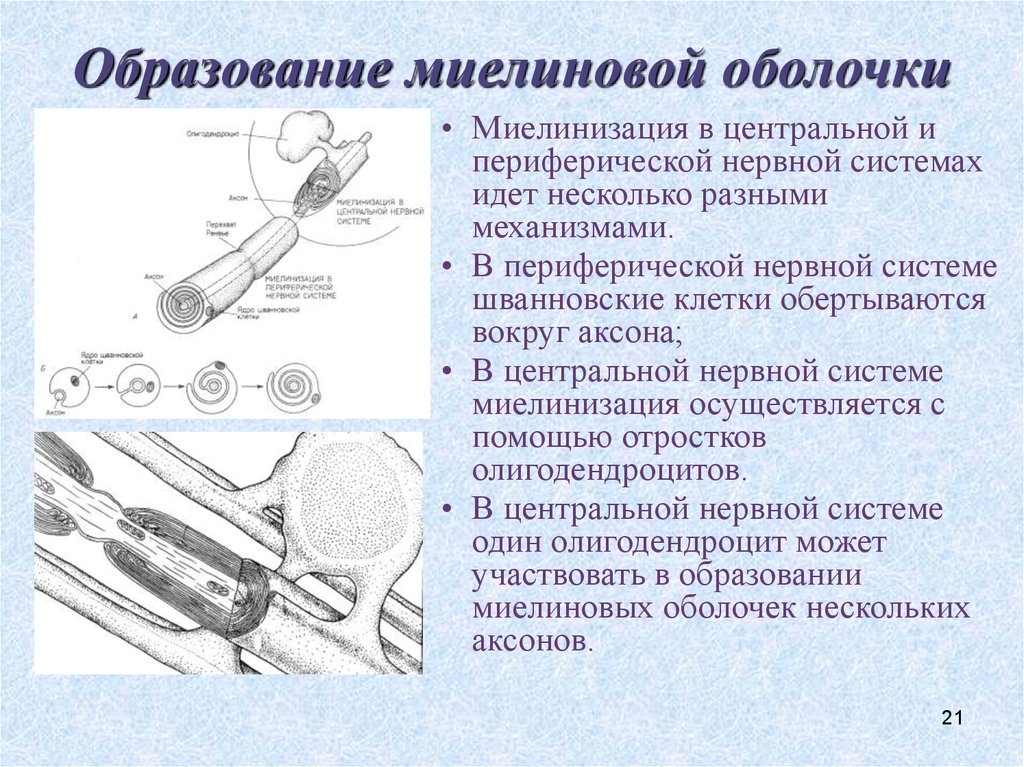
ОЖЖ және шеткері жүйке жүйесі (ШЖЖ) миелиндік қабығының түзілуі және миелин құрылымында айырмашылықтар бар. ОЖЖ миелині қалыптасуы кезінде
 бір олигодендроглиоцит бірнеше аксондардың миелинінің бірнеше сегменттерімен байланысады; бұл кезде аксонға олигодендроглиоциттің өсіндісі іргелесіп бекиді, бұл олигодендроглиоцит аксоннан едәуір қашықтықта орналасқан, миелиннің сыртқы беткейі жасушасыртылық кеңістікпен жанасады.
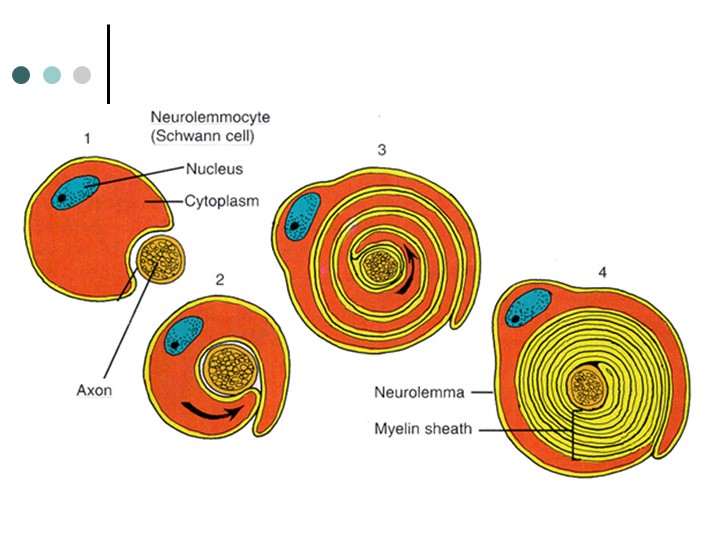
бір олигодендроглиоцит бірнеше аксондардың миелинінің бірнеше сегменттерімен байланысады; бұл кезде аксонға олигодендроглиоциттің өсіндісі іргелесіп бекиді, бұл олигодендроглиоцит аксоннан едәуір қашықтықта орналасқан, миелиннің сыртқы беткейі жасушасыртылық кеңістікпен жанасады.Шванн жасушасы ШЖЖ миелинінің түзілуі кезінде миелиннің ширатылған тақтайшаларын қалыптастырады және Ранвье қалтарыстарының арасындағы миелин қабықтарының жекелеген бөлімі үшін ғана жауап береді. Шванн жасушасының цитоплазмасы ширатпа иірімдері арасындағы кеңістіктен ығыстырылып шығарылады да, миелин қабығының сыртқы және ішкі беткейлерінде ғана қалады.
Миелиннің протеиндері: сипаттамасы
Миелин протеиндері ішкі (intrinsic) және сыртқы (extrinsic) протеиндер болып бөлінеді. Біріншілері мембрана арқылы өтеді және онымен берік байланысқан. Сыртқы протеиндер мембрана беткейінде әлсіз байланыса орналасқан. Мұндай мембрана химиялық құрамы және электр заряды бойынша асимметриялы болады. Оның экстрацеллюлярлық беткейі гликопротеиндер мен гликолипидтердің көмірсулы қалдықтарына бай, гликопротеиндердің С-соңы мембрананың цитоплазматикалық жағында, ал полисахаридтік қалдықтары экстрацеллюлярлық беткейінде орналасады. Миелин мембранасындағы липидтердің әртүрлі бағыттағы гидрофилді топтарының арақатынасы 4,5-5,0 нм, осы уақыттағы ширатпаның көршілес иірімдері арақашықтығы 3,0-5,0 нм құрайды. Липидтерден ғана қалыптастырылған мультиламеллярлық түзілістің қалыңдығы 1,5-3,0 нм болады. Миелиннің 29 протеиндері сипатталған.
 Басқа биологиялық мембраналардан миелиннің айырмашылығын көрсететін биохимиялық сипаттамалардың бірі липид/протеин жоғарғы қатынасы болып табылады. Протеиндер миелин қабығының құрғақ заттары массасының 25 - 30% құрайды. Липидтер үлесіне сүтқоректілердің ОЖЖ ақ затының құрғақ массасының ≈70-75% тиесілі; жұлынның миелинінде липид/протеин қатынасы жоғары болады. Липидтердің жалпы мөлшерінің 28% - холестерол, 43% – фосфолипидтер және 29% - галактолипидтер құрайды. Липидтер миелин мембранасы құрамына кіретін протеиндердің конформациялық сипаттарына зор әсер көрсетеді, ал протеиндер де өз кезегінде липидтердің қасиеттеріне әсерін тигізеді.
Басқа биологиялық мембраналардан миелиннің айырмашылығын көрсететін биохимиялық сипаттамалардың бірі липид/протеин жоғарғы қатынасы болып табылады. Протеиндер миелин қабығының құрғақ заттары массасының 25 - 30% құрайды. Липидтер үлесіне сүтқоректілердің ОЖЖ ақ затының құрғақ массасының ≈70-75% тиесілі; жұлынның миелинінде липид/протеин қатынасы жоғары болады. Липидтердің жалпы мөлшерінің 28% - холестерол, 43% – фосфолипидтер және 29% - галактолипидтер құрайды. Липидтер миелин мембранасы құрамына кіретін протеиндердің конформациялық сипаттарына зор әсер көрсетеді, ал протеиндер де өз кезегінде липидтердің қасиеттеріне әсерін тигізеді.
 Миелиннің негізгі қызметі аксондар бойынша жүйке импульсін тез өткізу болып табылады. Миелинді қалыптастыратын жасушалар мембраналары тығыз жанасады және жоғарғы кедергі мен төменгі сыйымдылықты қамтамасыз етеді, сөйтіп, аксонды тиімді оқшаулайды, импульстың бойлық таралуына жол бермейді. Миелин 1 мм жуық ұзындық аралығында кездесетін Ранвье қалтарыстары аймағында ғана үзіледі. Сондықтан иондар ағыны миелин арқылы өте алмайды, иондар алмасуы қалтарыстар аймағында ғана жүзеге асырылады. Бұл жүйке импульсының таралуы жылдамдығының жоғарылауына алып келеді. Сонымен, миелинделмегенге қарағанда, миелинделген талшықтар арқылы импульс 5-10 есе жылдам өтеді. Миелин жүйке импульсын таратудан басқа, жүйке талшықтарын қоректендіруге қатысады, құрылымдық және қорғаныстық қызметтерді де атқарады.
Миелиннің негізгі қызметі аксондар бойынша жүйке импульсін тез өткізу болып табылады. Миелинді қалыптастыратын жасушалар мембраналары тығыз жанасады және жоғарғы кедергі мен төменгі сыйымдылықты қамтамасыз етеді, сөйтіп, аксонды тиімді оқшаулайды, импульстың бойлық таралуына жол бермейді. Миелин 1 мм жуық ұзындық аралығында кездесетін Ранвье қалтарыстары аймағында ғана үзіледі. Сондықтан иондар ағыны миелин арқылы өте алмайды, иондар алмасуы қалтарыстар аймағында ғана жүзеге асырылады. Бұл жүйке импульсының таралуы жылдамдығының жоғарылауына алып келеді. Сонымен, миелинделмегенге қарағанда, миелинделген талшықтар арқылы импульс 5-10 есе жылдам өтеді. Миелин жүйке импульсын таратудан басқа, жүйке талшықтарын қоректендіруге қатысады, құрылымдық және қорғаныстық қызметтерді де атқарады. МИЕЛИННІҢ НЕГІЗГІ ПРОТЕИНІ
Құрылысы, физ.-хим. қасиеттері.
Негізгі массасы – барлық протеиндердің– протеолипидтік протеиндер (PLP)- М 30 кДа - 50% жуығын құрайды.
Протеиндер - миелиннің негізгі протеиндері тобы (МНП) -шамамен 30% М 17-21,5 кДа, бұл МНП 3 изоформасы.
Олардың біріншісі 7 экзонмен кодталады; 2 экзонның бөлінуі нәтижесінде протеин М 18,5 кДа синтезделеді, массасы 17,5 кДа болатын изоформасының кодталуы кезінде 5 экзон делецияға ұшырайды. Адамның МНП гені 18 хромосомада орналасқан және ақпараттың оқылуы басталатын 3 промоторлық аймағы болады. МНП құрамында (шамамен 25%) негізгі аминқышқылдары (аргинин, лизин и гистидин) бар, олар полипептидтік тізбекте біркелкі таралып орналасқан және МНП-нің өте жоғарғы изоэлектрилік нүктесін қамтамасыз етеді (рI=12-13).
МНП молекуласында аргинин цитруллинмен алмасқанда оның иммуногендік қабілеті жоғарылайды, бұл шашыраңқы склероз кезінде байқалады.
МНП фосфорлануы. Фосфорлану процесі гетерогендіктің негізгі шығу көзі болып табылады. Фосфорланған МНП гидрофилді болады және оң зарядтың мәні кемиді, сондықтан аниондық фосфолипидтермен байланысы әлсірейді.
Миелиннің протеинкиназалық жән фосфорилазалық белсенділігі болады, сондықтан ол МНП-нің шамадан тыс фосфорлануын реттейді. Миелиннің протеинкиназасы кальциймен, цАМФ және фосфатидилинозитол-4-фосфатмен белсендіріледі. Миелиногенез олигодендроциттің субстратқа адгезиясы кезінде жүзеге асырылады және МНП –нің протеинкиназа С көмегімен фосфорлануы арқылы күшейтіледі. Ганглиозидтер протеинкиназа С белсенділігін күшейтеді және протеинкиназа А белсенділігін тежейді. Ганглиозидтер, сонымен қатар, протеинкиназа С ферментінің диацилглицеролмен белсендірілуін басады. МНП –нің кейбір бөліктері протеинкиназа С ферменті белсенділігінің арнайы ингибиторы болып табылады.
МНП метилденуі. МНП басқа посттранскрипциялық түрленуі - 107орындағы аргинин қалдығының аргинин-метилтрансферазамен метилденуі. Бұл ферменттің экспрессиясы тиреоидтық гормондармен реттеледі. S-аденозил-L-метионин МНП құрамындағы аргинин қалдығы үшін метил тобының доноры болып табылады.
МНП метилдену дәрежесі даму процесінде едәуір өзгереді. Демиелинизацияның инициациясы S-аденозил-L-метионин биосинтезінің циклолейцинмен тежелуі кезінде мүмкін болады, миелиннің тығыз құрылымының қалыптасуын синефунгин –метилтрансферазаның ингибиторы басады.
МНП жоқтығы ШЖЖ тығыз миелинінің қалыптасуына кедергі келтірмейді, себебі миелиннің басқа протеині – Pо МНП-нің орнын толтырады. МНП –нің барлық изоформаларының N-соңы ацетилденеді, ал С-соңы карбоксипептидазалар әсеріне ұшырайды. МНП-не әртүрлі протеолиттік ферменттер әсер ете алады.. Катепсин D әсерінен протеиндердің гидролизденуі қышқылдық липидтер қатысқанда жылдамдайды.
МНП-нің минорлық фракциялары құрамында 54 орындағы серин қалдығы аймағымен ковалентті байланысқан фосфатидилинозитолбисфосфат болады.
МНП липидтермен өзара әрекеттесуі. МНП мембрана компоненті ретінде липидтермен жақсы байланыса алады. Сонымен қатар, поликатион болғандықтан МНП аниондық липидтермен тұрақты кешендер қалыптастырады.
Аниондық фосфолипидтердің басым бөлігімен МНП-і кешен қалыптастырған кезде гидрофобтық және электростатикалық әрекеттесулер маңызды рөл атқарады.
МНП орталық жүйке жүйесі олигодендроглияларының және миелиннің арнайы маркері болып табылатын GM4 ганглиозид құрамды көпқабатты липосомадан глюкозаның босап шығуын қамтамасыз етеді. МНП-ның тек қана фосфатидилхолин құрамды көпіршіктердің агрегациясын индуцирлеу қабілеті бар, алайда агрегация дәрежесі қышқылдық фосфолипидтер қатысқанда едәуір күшейеді. Алифаттық альдегидтер қатысқанда фосфолипидтік көпіршіктердің агрегациясы МНП әсерінен едәуір күшейеді. Алифаттық альдегидтер деңгейінің жоғарылауы демиелиндеуші аурулармен байланысты болуы мүмкін. Холестерол әсерінен МНП-нің фосфатидилсерин көпіршіктерінің агрегациясын стимулдеуші қабілеті артады.
МНП клиника-диагностикалық маңызы.
Шашыраңқы склероз диагностикасының маркері ретінде МНП –ді жұлын сұйықтығында анықтау патологиялық процестің белсенділігі жайындағы болжаушы критерий қызметін атқарады. РИА әдісімен МНП концентрациясы (и NSE және S-100) әртүрлі неврологиялық патологиясы бар балалар мен ересек адамдардың жұлын сұйықтығында анықталады: мысалы, шашыраңқы склероз (созылмалы-прогрессивті, рецидивті-ремиттеуші және осы екі типтің комбинациясы), цереброваскулярлық бұзылыстар ( ишемиялық және геморрагиялық инсульт), деменция (тамырлық және Альцгеймер түрі) және ОЖЖ инфекциялары. МНП ең жоғарғы деңгейі шашыраңқы склерозбен ауыратын науқастарда байқалған, сонымен қатар, МНП нақты жоғарылау деңгейі цереброваскулярлық инсульттар және балаларда энцефалопатия кезіндерінде анықталған.

