ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 27.04.2024
Просмотров: 58
Скачиваний: 0
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.

отолитовую мембрану. Среди волосков чувствительных клеток, так же как и в ампуллярных рецепторах, различают киноцилии и стереоцилии. Давление отолитов на волоски чувствительных клеток, а также смещение волосков при прямолинейных ускорениях является моментом трансформации механической энергии в электри-
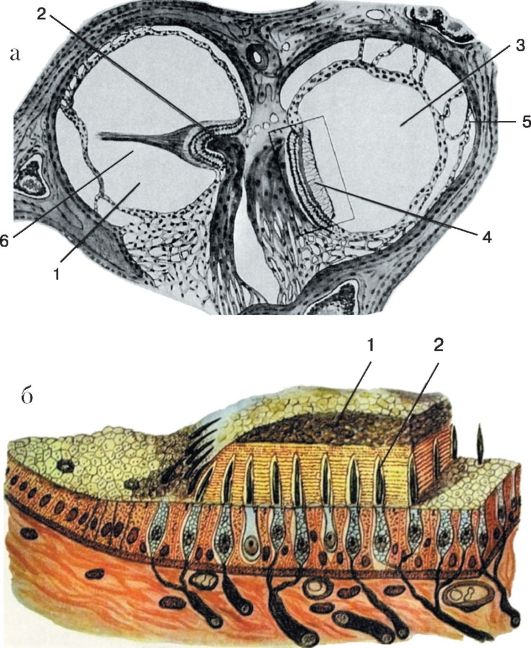 Рис. 5.15. Схема вестибулярных рецепторов:
Рис. 5.15. Схема вестибулярных рецепторов:
а - ампулярный рецептор: 1 - просвет ампулы полукружного протока; 2 - ампулярный гребешок; 3 - просвет эллиптического мешочка; 4 - мембрана статоконий; 5 - соединительнотканные тяжи; 6 - киноцилий; б - статокониев рецептор: 1 - мембрана статоконий; 2 - рецепторные клетки
ческую в нейроэпителиальных волосковых клетках. Эллиптический и сферический мешочки соединены между собой посредством тонкого канальца - ductus utriculosaccularis, который имеет ответвле-
ние - эндолимфатический проток (ductus endolimphaticus). Проходя в водопроводе преддверия, эндолимфатический проток выходит на заднюю поверхность пирамиды и там слепо заканчивается эндолимфатическим мешком (saccus endolimphaticus), представляющим собой расширение, образованное дупликатурой твердой мозговой оболочки.
Таким образом, вестибулярные сенсорные клетки расположены в пяти рецепторных областях: по одной в каждой ампуле трех полукружных каналов и по одной в двух мешочках преддверия каждого уха. В нервных рецепторах преддверия и полукружных каналов к каждой чувствительной клетке подходит не одно (как в улитке), а несколько нервных волокон, поэтому гибель одного из этих волокон не влечет за собой гибели клетки.
Кровоснабжение внутреннего уха осуществляется через лабиринтную артерию (a. labyrinthi), являющуюся ветвью базилярной артерии (a. basilaris) или ее ветви от передней нижней мозжечковой артерии (рис. 5.16). Во внутреннем слуховом проходе лабиринтная артерия делится на три ветви: преддверную (a. vestibularis), преддверно-улитковую (a. vestibulocochlearis) и улитковую (a. cochlearis).
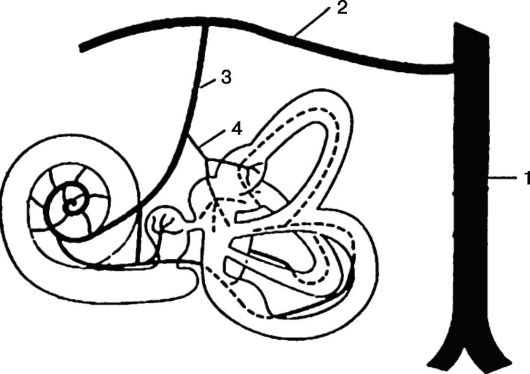 Рис. 5.16. Кровоснабжение лабиринта:
Рис. 5.16. Кровоснабжение лабиринта:
1 - позвоночная артерия; 2 - базилярная артерия; 3 - передняя нижняя мозжечковая артерия; 4 - артерия лабиринта
Особенности кровоснабжения лабиринта состоят в том, что ветви лабиринтной артерии не имеют анастомозов с сосудистой системой среднего уха, рейсснерова мембрана лишена капилляров, а в области ампулярных и отолитовых рецепторов подэпителиальная капиллярная сеть находится в непосредственном контакте с клетками нейроэпителия. К нейроэпителиальным волосковым клеткам спирального органа кровеносные сосуды не подходят, их питание осуществляется опосредованно через прилежащие к ним трофические клетки.
Венозный отток из внутреннего уха идет по трем путям: венам водопровода улитки, венам водопровода преддверия и венам внутреннего слухового прохода.
ИННЕРВАЦИЯ ВНУТРЕННЕГО УХА
Слуховой анализатор (рис. 5.17). Волосковые клетки кортиева органа синаптически связаны с периферическими отростками биполярных клеток спирального ганглия (ganglion spirale), расположенного в основании спиральной пластинки улитки. Центральные отростки биполярных нейронов спирального ганглия являются волокнами слуховой (улитковой) порции VIII нерва (n. cochleovestibularis), который проходит через внутренний слуховой проход и в области мостомозжечкового угла входит в мост. На дне четвертого желудочка VIII нерв делится на два корешка: верхний вестибулярный и нижний улитковый.
Волокна улиткового корешка заканчиваются в латеральном углу ромбовидной ямки на клетках вентрального ядра (nucl. ventralis) и дорсального улиткового ядра (nucl. dorsalis). Таким образом, клетки спирального ганглия вместе с периферическими отростками, идущими к нейроэпителиальным волосковым клеткам органа Корти, и центральными отростками, заканчивающимися в ядрах моста, составляют I нейрон слухового анализатора. На уровне кохлеарных ядер расположен ряд ядерных образований, принимающих участие в формировании дальнейших путей для проведения слуховых раздражений: ядро трапециевидного тела, верхняя олива, ядро боковой петли. От вентрального и дорсального ядер начинается II нейрон слухового анализатора. Меньшая часть волокон этого нейрона идет по одноименной стороне, a большая часть в виде striae acusticae перекрещиваются и переходят на противоположную сторону моста, заканчиваясь в оливе и трапециевидном теле. Волокна

III нейрона в составе боковой петли идут к ядрам четверохолмия и медиально-
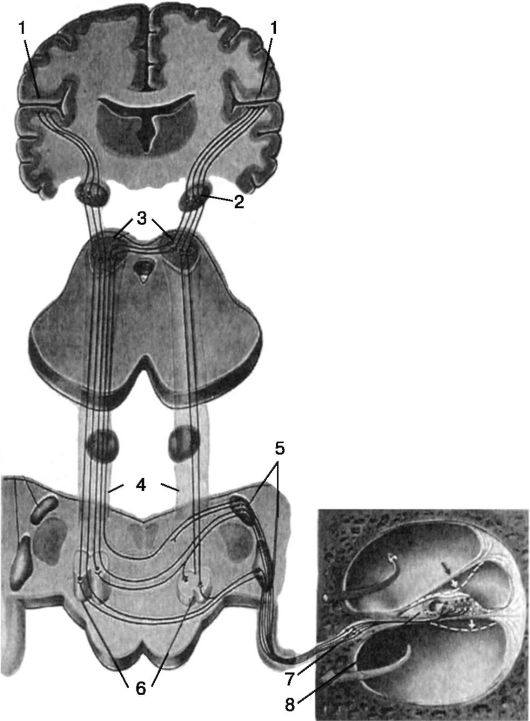 Рис. 5.17. Схема проводящих путей слухового анализатора: 1 - кора височной доли большого мозга; 2 - медиальное коленчатое тело; 3 - бугры четверохолмия; 4 - латеральная петля; 5 - улитковые ядра; 6 -верхние оливные ядра; 7 - спиральный узел; 8 - кортиев орган
Рис. 5.17. Схема проводящих путей слухового анализатора: 1 - кора височной доли большого мозга; 2 - медиальное коленчатое тело; 3 - бугры четверохолмия; 4 - латеральная петля; 5 - улитковые ядра; 6 -верхние оливные ядра; 7 - спиральный узел; 8 - кортиев орган
го коленчатого тела, откуда уже волокна IV нейрона после второго частичного перекреста направляются в височную долю мозга и оканчиваются в корковом отделе слухового анализатора, располагаясь преимущественно в поперечных височных извилинах Гешля.
Проведение импульсов от кохлеарных рецепторов по обеим сторонам мозгового ствола объясняет то обстоятельство, что односторон-
нее нарушение слуха возникает только в случае поражения среднего и внутреннего уха, а также кохлеовестибулярного нерва и его ядер в мосту. При одностороннем поражении латеральной петли, подкорковых и корковых слуховых центров импульсы от обоих кохлеарных рецепторов проводятся по непораженной стороне в одно из полушарий и расстройства слуха может не быть.
Слуховая система обеспечивает восприятие звуковых колебаний, проведение нервных импульсов к слуховым нервным центрам, анализ получаемой информации.
Вестибулярный анализатор. Рецепторные клетки вестибулярного анализатора контактируют с окончаниями периферических отростков биполярных нейронов вестибулярного ганглия (gangl. vestibulare), расположенного во внутреннем слуховом проходе. Центральные отростки этих нейронов формируют вестибулярную порцию преддверно-улиткового (VIII) нерва, который проходит во внутреннем слуховом проходе, выходит в заднюю черепную ямку и в области мостомозжечкового угла внедряется в вещество мозга. В вестибулярных ядрах продолговатого мозга, в дне четвертого желудочка, заканчивается I нейрон. Вестибулярный ядерный комплекс включает четыре ядра: латеральное, медиальное, верхнее и нисходящее. От каждого ядра идет с преимущественным перекрестом II нейрон.
Высокие адаптационные возможности вестибулярного анализатора обусловлены наличием множества ассоциативных путей ядерного вестибулярного комплекса (рис. 5.18). С позиций клинической анатомии важно отметить пять основных связей вестибулярных ядер с различными образованиями центральной и периферической нервной системы.
*Вестибулоспинальные связи. Начинаясь от латеральных ядер продолговатого мозга, в составе вестибулоспинального тракта, они проходят в передних рогах спинного мозга, обеспечивая связь вестибулярных рецепторов с мышечной системой. *Вестибулоглазодвигательные связи осуществляются через систему заднего продольного пучка: от медиального и нисходящего ядер продолговатого мозга идет перекрещенный путь, а от верхнего ядра - неперекрещенный, к глазодвигательным ядрам. *Вестибуловегетативные связи осуществляются от медиального ядра к ядрам блуждающего нерва, ретикулярной фармации, диэнцефальной области.
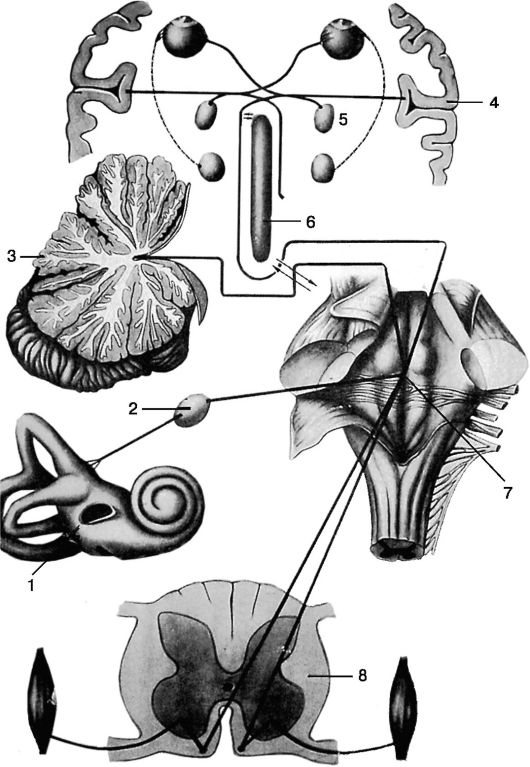 Рис. 5.18. Схема ассоциативных связей вестибулярного анализатора: 1 - лабиринт; 2 - спиральный ганглий; 3 - мозжечок; 4 - кора полушарий большого мозга; 5 - ядра глазодвигательных нервов; 6 - ретикулярная формация; 7 - вестибулярные ядра в продолговатом мозге; 8 - спинной мозг
Рис. 5.18. Схема ассоциативных связей вестибулярного анализатора: 1 - лабиринт; 2 - спиральный ганглий; 3 - мозжечок; 4 - кора полушарий большого мозга; 5 - ядра глазодвигательных нервов; 6 - ретикулярная формация; 7 - вестибулярные ядра в продолговатом мозге; 8 - спинной мозг
*Вестибуломозжечковые пути проходят во внутреннем отделе нижней ножки мозжечка и связывают вестибулярные ядра с ядрами мозжечка.
*Вестибулокортикальные связи обеспечиваются системой волокон, идущих от всех четырех ядер к зрительному бугру. Прерываясь в последнем, далее эти волокна идут к височной доле мозга, где вестибулярный анализатор имеет рассеянное представительство. Кора и мозжечок выполняют регулирующую функцию по отношению к вестибулярному анализатору.
Посредством указанных связей реализуются разнообразные сенсорные, вегетативные и соматические вестибулярные реакции.
КЛИНИЧЕСКАЯ ФИЗИОЛОГИЯ УХА
В ухе расположены в одной костной капсуле рецепторы двух органов (анализаторов) - слуха и равновесия. Оба они относятся к механорецепторам и характеризуются определенной сходностью восприятия энергии раздражения. В то же время более молодой в филогенетическом и онтогенетическом отношениях слуховой аппарат уха отличается большей сложностью организации. Чувствительные элементы слухового анализатора, в отличие от вестибулярных, относятся к экстероцепторам, т.е. воспринимают энергию из внешней среды. Рассмотрим особенности функционирования органов слуха и равновесия.
ФУНКЦИЯ ОРГАНА СЛУХА
Слух человека является сложным процессом, для реализации которого необходимо проведение звуковой волны, преобразование ее в электрические нервные импульсы, передача их в нервные центры, анализ и интеграция звуковой информации. Соответственно различают такие функции органа слуха, как звукопроведение и звуковосприятие. Адекватным раздражителем органа слуха является звук, поэтому для освещения основных функциональных особенностей системы необходимо знакомство с некоторыми понятиями акустики.
Основные физические понятия акустики. Звук представляет собой механические колебания упругой среды, распространяющиеся в виде волн в воздухе, жидкости и твердых телах. Источником звука может быть любой процесс, вызывающий местное изменение давле-
ния или механическое напряжение в среде. С точки зрения физиологии под звуком понимают такие механические колебания, которые, воздействуя на слуховой рецептор, вызывают в нем определенный физиологический процесс, воспринимаемый как ощущение звука.
Звуковая волна характеризуется синусоидальными, т.е. периодическими колебаниями (рис. 5.19). При распространении в определенной среде звук представляет собой волну с фазами сгущения (уплотнения) и разрежения. Различают волны поперечные - в твердых телах, и продольные - в воздухе и жидких средах. Скорость распространения звуковых колебаний в воздухе составляет 332 м/с, в воде - 1450 м/с. Одинаковые состояния звуковой волны - участки сгущения или разрежения - называются
Рис. 5.15. Схема вестибулярных рецепторов:а - ампулярный рецептор: 1 - просвет ампулы полукружного протока; 2 - ампулярный гребешок; 3 - просвет эллиптического мешочка; 4 - мембрана статоконий; 5 - соединительнотканные тяжи; 6 - киноцилий; б - статокониев рецептор: 1 - мембрана статоконий; 2 - рецепторные клетки
ческую в нейроэпителиальных волосковых клетках. Эллиптический и сферический мешочки соединены между собой посредством тонкого канальца - ductus utriculosaccularis, который имеет ответвле-
ние - эндолимфатический проток (ductus endolimphaticus). Проходя в водопроводе преддверия, эндолимфатический проток выходит на заднюю поверхность пирамиды и там слепо заканчивается эндолимфатическим мешком (saccus endolimphaticus), представляющим собой расширение, образованное дупликатурой твердой мозговой оболочки.
| |
Таким образом, вестибулярные сенсорные клетки расположены в пяти рецепторных областях: по одной в каждой ампуле трех полукружных каналов и по одной в двух мешочках преддверия каждого уха. В нервных рецепторах преддверия и полукружных каналов к каждой чувствительной клетке подходит не одно (как в улитке), а несколько нервных волокон, поэтому гибель одного из этих волокон не влечет за собой гибели клетки.
Кровоснабжение внутреннего уха осуществляется через лабиринтную артерию (a. labyrinthi), являющуюся ветвью базилярной артерии (a. basilaris) или ее ветви от передней нижней мозжечковой артерии (рис. 5.16). Во внутреннем слуховом проходе лабиринтная артерия делится на три ветви: преддверную (a. vestibularis), преддверно-улитковую (a. vestibulocochlearis) и улитковую (a. cochlearis).
Рис. 5.16. Кровоснабжение лабиринта:1 - позвоночная артерия; 2 - базилярная артерия; 3 - передняя нижняя мозжечковая артерия; 4 - артерия лабиринта
Особенности кровоснабжения лабиринта состоят в том, что ветви лабиринтной артерии не имеют анастомозов с сосудистой системой среднего уха, рейсснерова мембрана лишена капилляров, а в области ампулярных и отолитовых рецепторов подэпителиальная капиллярная сеть находится в непосредственном контакте с клетками нейроэпителия. К нейроэпителиальным волосковым клеткам спирального органа кровеносные сосуды не подходят, их питание осуществляется опосредованно через прилежащие к ним трофические клетки.
Венозный отток из внутреннего уха идет по трем путям: венам водопровода улитки, венам водопровода преддверия и венам внутреннего слухового прохода.
ИННЕРВАЦИЯ ВНУТРЕННЕГО УХА
Слуховой анализатор (рис. 5.17). Волосковые клетки кортиева органа синаптически связаны с периферическими отростками биполярных клеток спирального ганглия (ganglion spirale), расположенного в основании спиральной пластинки улитки. Центральные отростки биполярных нейронов спирального ганглия являются волокнами слуховой (улитковой) порции VIII нерва (n. cochleovestibularis), который проходит через внутренний слуховой проход и в области мостомозжечкового угла входит в мост. На дне четвертого желудочка VIII нерв делится на два корешка: верхний вестибулярный и нижний улитковый.
| |
Волокна улиткового корешка заканчиваются в латеральном углу ромбовидной ямки на клетках вентрального ядра (nucl. ventralis) и дорсального улиткового ядра (nucl. dorsalis). Таким образом, клетки спирального ганглия вместе с периферическими отростками, идущими к нейроэпителиальным волосковым клеткам органа Корти, и центральными отростками, заканчивающимися в ядрах моста, составляют I нейрон слухового анализатора. На уровне кохлеарных ядер расположен ряд ядерных образований, принимающих участие в формировании дальнейших путей для проведения слуховых раздражений: ядро трапециевидного тела, верхняя олива, ядро боковой петли. От вентрального и дорсального ядер начинается II нейрон слухового анализатора. Меньшая часть волокон этого нейрона идет по одноименной стороне, a большая часть в виде striae acusticae перекрещиваются и переходят на противоположную сторону моста, заканчиваясь в оливе и трапециевидном теле. Волокна
III нейрона в составе боковой петли идут к ядрам четверохолмия и медиально-
Рис. 5.17. Схема проводящих путей слухового анализатора: 1 - кора височной доли большого мозга; 2 - медиальное коленчатое тело; 3 - бугры четверохолмия; 4 - латеральная петля; 5 - улитковые ядра; 6 -верхние оливные ядра; 7 - спиральный узел; 8 - кортиев органго коленчатого тела, откуда уже волокна IV нейрона после второго частичного перекреста направляются в височную долю мозга и оканчиваются в корковом отделе слухового анализатора, располагаясь преимущественно в поперечных височных извилинах Гешля.
Проведение импульсов от кохлеарных рецепторов по обеим сторонам мозгового ствола объясняет то обстоятельство, что односторон-
нее нарушение слуха возникает только в случае поражения среднего и внутреннего уха, а также кохлеовестибулярного нерва и его ядер в мосту. При одностороннем поражении латеральной петли, подкорковых и корковых слуховых центров импульсы от обоих кохлеарных рецепторов проводятся по непораженной стороне в одно из полушарий и расстройства слуха может не быть.
| |
Слуховая система обеспечивает восприятие звуковых колебаний, проведение нервных импульсов к слуховым нервным центрам, анализ получаемой информации.
Вестибулярный анализатор. Рецепторные клетки вестибулярного анализатора контактируют с окончаниями периферических отростков биполярных нейронов вестибулярного ганглия (gangl. vestibulare), расположенного во внутреннем слуховом проходе. Центральные отростки этих нейронов формируют вестибулярную порцию преддверно-улиткового (VIII) нерва, который проходит во внутреннем слуховом проходе, выходит в заднюю черепную ямку и в области мостомозжечкового угла внедряется в вещество мозга. В вестибулярных ядрах продолговатого мозга, в дне четвертого желудочка, заканчивается I нейрон. Вестибулярный ядерный комплекс включает четыре ядра: латеральное, медиальное, верхнее и нисходящее. От каждого ядра идет с преимущественным перекрестом II нейрон.
Высокие адаптационные возможности вестибулярного анализатора обусловлены наличием множества ассоциативных путей ядерного вестибулярного комплекса (рис. 5.18). С позиций клинической анатомии важно отметить пять основных связей вестибулярных ядер с различными образованиями центральной и периферической нервной системы.
*Вестибулоспинальные связи. Начинаясь от латеральных ядер продолговатого мозга, в составе вестибулоспинального тракта, они проходят в передних рогах спинного мозга, обеспечивая связь вестибулярных рецепторов с мышечной системой. *Вестибулоглазодвигательные связи осуществляются через систему заднего продольного пучка: от медиального и нисходящего ядер продолговатого мозга идет перекрещенный путь, а от верхнего ядра - неперекрещенный, к глазодвигательным ядрам. *Вестибуловегетативные связи осуществляются от медиального ядра к ядрам блуждающего нерва, ретикулярной фармации, диэнцефальной области.
| |
Рис. 5.18. Схема ассоциативных связей вестибулярного анализатора: 1 - лабиринт; 2 - спиральный ганглий; 3 - мозжечок; 4 - кора полушарий большого мозга; 5 - ядра глазодвигательных нервов; 6 - ретикулярная формация; 7 - вестибулярные ядра в продолговатом мозге; 8 - спинной мозг*Вестибуломозжечковые пути проходят во внутреннем отделе нижней ножки мозжечка и связывают вестибулярные ядра с ядрами мозжечка.
*Вестибулокортикальные связи обеспечиваются системой волокон, идущих от всех четырех ядер к зрительному бугру. Прерываясь в последнем, далее эти волокна идут к височной доле мозга, где вестибулярный анализатор имеет рассеянное представительство. Кора и мозжечок выполняют регулирующую функцию по отношению к вестибулярному анализатору.
Посредством указанных связей реализуются разнообразные сенсорные, вегетативные и соматические вестибулярные реакции.
КЛИНИЧЕСКАЯ ФИЗИОЛОГИЯ УХА
В ухе расположены в одной костной капсуле рецепторы двух органов (анализаторов) - слуха и равновесия. Оба они относятся к механорецепторам и характеризуются определенной сходностью восприятия энергии раздражения. В то же время более молодой в филогенетическом и онтогенетическом отношениях слуховой аппарат уха отличается большей сложностью организации. Чувствительные элементы слухового анализатора, в отличие от вестибулярных, относятся к экстероцепторам, т.е. воспринимают энергию из внешней среды. Рассмотрим особенности функционирования органов слуха и равновесия.
ФУНКЦИЯ ОРГАНА СЛУХА
Слух человека является сложным процессом, для реализации которого необходимо проведение звуковой волны, преобразование ее в электрические нервные импульсы, передача их в нервные центры, анализ и интеграция звуковой информации. Соответственно различают такие функции органа слуха, как звукопроведение и звуковосприятие. Адекватным раздражителем органа слуха является звук, поэтому для освещения основных функциональных особенностей системы необходимо знакомство с некоторыми понятиями акустики.
| |
Основные физические понятия акустики. Звук представляет собой механические колебания упругой среды, распространяющиеся в виде волн в воздухе, жидкости и твердых телах. Источником звука может быть любой процесс, вызывающий местное изменение давле-
ния или механическое напряжение в среде. С точки зрения физиологии под звуком понимают такие механические колебания, которые, воздействуя на слуховой рецептор, вызывают в нем определенный физиологический процесс, воспринимаемый как ощущение звука.
Звуковая волна характеризуется синусоидальными, т.е. периодическими колебаниями (рис. 5.19). При распространении в определенной среде звук представляет собой волну с фазами сгущения (уплотнения) и разрежения. Различают волны поперечные - в твердых телах, и продольные - в воздухе и жидких средах. Скорость распространения звуковых колебаний в воздухе составляет 332 м/с, в воде - 1450 м/с. Одинаковые состояния звуковой волны - участки сгущения или разрежения - называются