Файл: 1. Гистология как наука. Ее взаимосвязь с другими дисциплинами и роль в формирование врача ветеринарной медицины. Гистология.docx
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 18.10.2024
Просмотров: 9
Скачиваний: 0
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.

мембран. Такая общность строения объясняется тем, что роль митохондрии одинакова: осуществление определенных ферментных реакций.
При исследовании «вглубь» по-иному предстало и основное вещество цитоплазмы клеток. В световом микроскопе оно выглядело по-разному. Дело в том, что живая клетка при изучении обычно фиксируется — убивается. При этом внутреннее строение ее в той или иной степени нарушается: иногда становится бесструктурным, иногда грубозернистым, нередко заполняется массой пузырьков, так как происходит свертывание белков.
Совсем иную картину дает электронный микроскоп: перед нами целая сеть нитей, трубочек, пузырьков. Все они ограничены тончайшими (примерно такими же, как у митохондрий) мембранами, часто усеянными мелкими зернышками. Эти структуры, получившие название эргастоплазменной сети, впервые представ перед исследователями, вызвали массу споров. Многие не верили в их реальность: настолько это было ново и неожиданно. Сейчас дискуссии постепенно затихают. Такие сети обнаружены почти во всех клетках. Начинает проясняться их важная роль. Установлена связь эргастоплазменной сети с особыми участками клетки — базофильными структурами.
Связь осуществляется через мелкие зернышки, усеивающие мембраны эргастоплазмы. Эти зернышки содержат одно из важнейших веществ клетки — рибонуклеиновую кислоту, которая играет активную роль в синтезе белка. Действительно, наиболее значительное скопление эргастоплазменной сети обнаруживается как раз в тех клетках, которые вырабатывают белки (например, поджелудочная железа).
16. Характеристика клеточных включений.
Включения — это непостоянные структуры клетки, которые появляются в ней и исчезают в процессе метаболизма. Различают трофические, секреторные, экскреторные и пигментные включения.
Группа трофических включений объединяет углеводные, липидные и белковые включения. Наиболее распространенным представителем углеводных включений является гликоген — полимер глюкозы. На светооптическом уровне наблюдать включения гликогена можно при использовании гистохимической ШИК-реакции. В электронном микроскопе гликоген выявляется как осмиофильные гранулы, которые в клетках, где гликогена много (гепатоцитах), сливаются в крупные конгломераты — глыбки. Липидными включениями наиболее богаты клетки жировой ткани — липоциты, резервирующие запасы жира для нужд всего организма, а также стероидпродуцирующие эндокринные клетки, использующие липид холестерин для синтеза своих гормонов. На ультрамикроскопическом уровне липидные включения имеют правильную округлую форму и в зависимости от химического состава характеризуются высокой, средней или низкой электронной плотностью.
Белковые включения, например, вителлин в яйцеклетках, накапливается в цитоплазме в виде гранул.
Секреторные включения представляют собой разнообразную группу. Секреторные включения синтезируются в клетках и выделяются (секретируются) в просветы протоков (клетки экзокринных желез), в межклеточную среду (гормоны, нейромедиаторы, факторы роста и др.), кровь, лимфу, межклеточные пространства (гормоны). На ультрамикроскопическом уровне секреторные включения имеют вид мембранных пузырьков, содержащих вещества разной плотности и интенсивности окраски, что зависит от их химического состава.
Экскреторные включения — это, как правило, продукты метаболизма клетки, от которых она должна освободиться. К экскреторным включениям относятся также инородные включения — случайно, либо преднамеренно (при фагоцитозе бактерий, например,) попавшие в клетку субстраты. Такие включения клетка лизирует с помощью своей лизосомальной системы, а оставшиеся частицы выводит (экскретирует ) во внешнюю среду. В более редких случаях попавшие в клетку агенты остаются неизменными и могут не подвергнуться экскреции — такие включения более правильно именовать чужеродными (хотя чужеродными для клетки являются и включения, которые она лизирует).
17. Различные типы секреции (апокриновый, мерокриновый, голокриновый). Участие органеллы в секреции.
18. Строение и функции ядра.
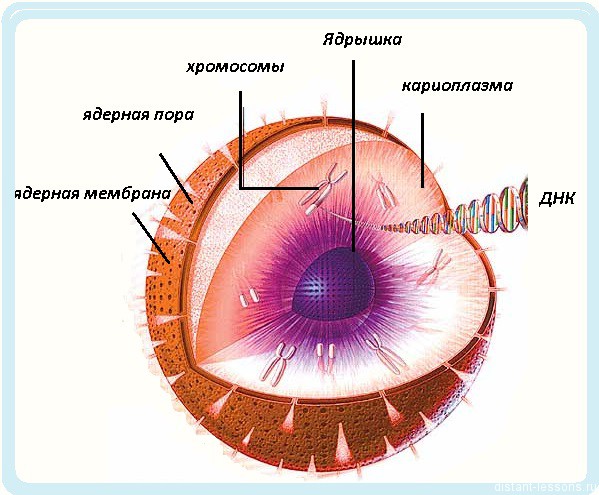
Как правило, эукариотическая клетка имеет одно ядро, но встречаются двуядерные (инфузории) и многоядерные клетки (опалина). Некоторые высокоспециализированные клетки вторично утрачивают ядро (эритроциты млекопитающих, ситовидные трубки покрытосеменных).
Форма ядра — сферическая, эллипсовидная, реже лопастная, бобовидная и др. Диаметр ядра — обычно от 3 до 10 мкм.
Строение ядра:
1 — наружная мембрана; 2 — внутренняя мембрана; 3 — поры; 4 — ядрышко; 5 — гетерохроматин; 6 — эухроматин.

Ядро отграничено от цитоплазмы двумя мембранами (каждая из них имеет типичное строение). Между мембранами — узкая щель, заполненная полужидким веществом. В некоторых местах мембраны сливаются друг с другом, образуя поры (3), через которые происходит обмен веществ между ядром и цитоплазмой.
Функции ядра:
1) хранение наследственной информации и передача ее дочерним клеткам в процессе деления,
2) регуляция жизнедеятельности клетки путем регуляции синтеза различных белков,
3) место образования субъединиц рибосом.
19. Виды клеточного деления.
Митоз – Не прямое деление. Образование дочерних клеток, генетически идентичных материнской. (рост, обновление, регенерация, бесполое размножение).
2. Мейоз – два последовательных деления, ведущие к уменьшению набора хромосом в 2 раза. При половом размножении обеспечивает постоянство набора хромосом и генетическую рекомбинацию в потомстве.
3. Эндомитоз – деление “внутри” клетки или ядра. Образуются двуядерные или полиплоидные клетки.
4. Патологические деления - амитоз, трехполюсной митоз и др.
20. Биологическая сущность и характеристика фаз митоза
Биологическое значение митоза — образование генетически одинаковых дочерних клеток с тем же набором хромосом, что был у материнской клетки. Сохраняется преемственность в ряду клеточных поколений.
фазы митоза:
При исследовании «вглубь» по-иному предстало и основное вещество цитоплазмы клеток. В световом микроскопе оно выглядело по-разному. Дело в том, что живая клетка при изучении обычно фиксируется — убивается. При этом внутреннее строение ее в той или иной степени нарушается: иногда становится бесструктурным, иногда грубозернистым, нередко заполняется массой пузырьков, так как происходит свертывание белков.
Совсем иную картину дает электронный микроскоп: перед нами целая сеть нитей, трубочек, пузырьков. Все они ограничены тончайшими (примерно такими же, как у митохондрий) мембранами, часто усеянными мелкими зернышками. Эти структуры, получившие название эргастоплазменной сети, впервые представ перед исследователями, вызвали массу споров. Многие не верили в их реальность: настолько это было ново и неожиданно. Сейчас дискуссии постепенно затихают. Такие сети обнаружены почти во всех клетках. Начинает проясняться их важная роль. Установлена связь эргастоплазменной сети с особыми участками клетки — базофильными структурами.
Связь осуществляется через мелкие зернышки, усеивающие мембраны эргастоплазмы. Эти зернышки содержат одно из важнейших веществ клетки — рибонуклеиновую кислоту, которая играет активную роль в синтезе белка. Действительно, наиболее значительное скопление эргастоплазменной сети обнаруживается как раз в тех клетках, которые вырабатывают белки (например, поджелудочная железа).
16. Характеристика клеточных включений.
Включения — это непостоянные структуры клетки, которые появляются в ней и исчезают в процессе метаболизма. Различают трофические, секреторные, экскреторные и пигментные включения.
Группа трофических включений объединяет углеводные, липидные и белковые включения. Наиболее распространенным представителем углеводных включений является гликоген — полимер глюкозы. На светооптическом уровне наблюдать включения гликогена можно при использовании гистохимической ШИК-реакции. В электронном микроскопе гликоген выявляется как осмиофильные гранулы, которые в клетках, где гликогена много (гепатоцитах), сливаются в крупные конгломераты — глыбки. Липидными включениями наиболее богаты клетки жировой ткани — липоциты, резервирующие запасы жира для нужд всего организма, а также стероидпродуцирующие эндокринные клетки, использующие липид холестерин для синтеза своих гормонов. На ультрамикроскопическом уровне липидные включения имеют правильную округлую форму и в зависимости от химического состава характеризуются высокой, средней или низкой электронной плотностью.
Белковые включения, например, вителлин в яйцеклетках, накапливается в цитоплазме в виде гранул.
Секреторные включения представляют собой разнообразную группу. Секреторные включения синтезируются в клетках и выделяются (секретируются) в просветы протоков (клетки экзокринных желез), в межклеточную среду (гормоны, нейромедиаторы, факторы роста и др.), кровь, лимфу, межклеточные пространства (гормоны). На ультрамикроскопическом уровне секреторные включения имеют вид мембранных пузырьков, содержащих вещества разной плотности и интенсивности окраски, что зависит от их химического состава.
Экскреторные включения — это, как правило, продукты метаболизма клетки, от которых она должна освободиться. К экскреторным включениям относятся также инородные включения — случайно, либо преднамеренно (при фагоцитозе бактерий, например,) попавшие в клетку субстраты. Такие включения клетка лизирует с помощью своей лизосомальной системы, а оставшиеся частицы выводит (экскретирует ) во внешнюю среду. В более редких случаях попавшие в клетку агенты остаются неизменными и могут не подвергнуться экскреции — такие включения более правильно именовать чужеродными (хотя чужеродными для клетки являются и включения, которые она лизирует).
17. Различные типы секреции (апокриновый, мерокриновый, голокриновый). Участие органеллы в секреции.
-
мерокриновые железы: секрет выделяется из клетки без нарушения ее целостности. К ним относятся большинство желез, например, слюнные железы; -
апокриновые железы: апикальная часть клетки отторгается вместе с секретом. К апокриновым железам относятся молочные и потовые железы; -
голокриновые железы: после накопления секрета клетка полностью разрушается и ее остатки включаются в состав секрета. К голокриновым железам относятся сальные железы кожи.
18. Строение и функции ядра.
Как правило, эукариотическая клетка имеет одно ядро, но встречаются двуядерные (инфузории) и многоядерные клетки (опалина). Некоторые высокоспециализированные клетки вторично утрачивают ядро (эритроциты млекопитающих, ситовидные трубки покрытосеменных).
Форма ядра — сферическая, эллипсовидная, реже лопастная, бобовидная и др. Диаметр ядра — обычно от 3 до 10 мкм.
Строение ядра:
1 — наружная мембрана; 2 — внутренняя мембрана; 3 — поры; 4 — ядрышко; 5 — гетерохроматин; 6 — эухроматин.
Ядро отграничено от цитоплазмы двумя мембранами (каждая из них имеет типичное строение). Между мембранами — узкая щель, заполненная полужидким веществом. В некоторых местах мембраны сливаются друг с другом, образуя поры (3), через которые происходит обмен веществ между ядром и цитоплазмой.
Функции ядра:
1) хранение наследственной информации и передача ее дочерним клеткам в процессе деления,
2) регуляция жизнедеятельности клетки путем регуляции синтеза различных белков,
3) место образования субъединиц рибосом.
19. Виды клеточного деления.
Митоз – Не прямое деление. Образование дочерних клеток, генетически идентичных материнской. (рост, обновление, регенерация, бесполое размножение).
2. Мейоз – два последовательных деления, ведущие к уменьшению набора хромосом в 2 раза. При половом размножении обеспечивает постоянство набора хромосом и генетическую рекомбинацию в потомстве.
3. Эндомитоз – деление “внутри” клетки или ядра. Образуются двуядерные или полиплоидные клетки.
4. Патологические деления - амитоз, трехполюсной митоз и др.
20. Биологическая сущность и характеристика фаз митоза
Биологическое значение митоза — образование генетически одинаковых дочерних клеток с тем же набором хромосом, что был у материнской клетки. Сохраняется преемственность в ряду клеточных поколений.
фазы митоза:
-
Профаза (2n4с) — спирализация хромосом, уменьшение их функциональной активности; репликация практически не идёт; разрушение оболочки ядра; образование веретена деления. -
Метафаза (2n4с) — прикрепление хромосом к нитям веретена деления; спирализация хромосом достигает максимума; хромосомы утрачивают свою функциональную активность, образуют экваториальную (метафазную) пластинку. -
Анафаза (4n4c) — деление центромер; расхождение по нитям веретена сестринских хромосом. Анафаза заканчивается, когда центромеры достигают полюсов клетки. -
Телофаза (2n2c) — деспирализация хромосом; образование ядерной оболочки; деление цитоплазмы; между дочерними клетками на экваторе образуется перетяжка. В растительных и грибных клетках в этом месте начинает закладываться клеточная стенка.