Файл: Гистологическая картина крови, при железодефицитной анемии.docx
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 12.04.2024
Просмотров: 13
Скачиваний: 0
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.

Анемия
| Малокровие | |
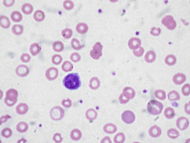 | |
| Гистологическая картина крови, при железодефицитной анемии | |
| | |
| | |
| | |
| | |
Анемия (греч. αναιμία, малокровие) — группа клинико-гематологических синдромов, общим моментом для которых является снижение концентрации гемоглобина в крови, чаще при одновременном уменьшении числа эритроцитов (или общего объёма эритроцитов). Термин «анемия» без детализации не определяет конкретного заболевания, то есть анемию следует считать одним из симптомов различных патологических состояний. Следует различать гидремию и анемию, при гидремии число форменных элементов и гемоглобина остается прежним, но увеличивается объём жидкой части крови.
Особенности классификации
Сама по себе любая анемия не является заболеванием, но может встречаться как синдром при целом ряде заболеваний, которые могут быть либо связаны с первичным поражением системы крови, либо не зависеть от него. В связи с этим строгая нозологическая классификация анемий невозможна. Для классификации анемий принято использовать принцип практической целесообразности. Для этого наиболее удобно делить анемии по единому классификационному признаку — цветовому показателю.
Снижение концентрации гемоглобина в крови часто происходит при одновременном уменьшении количества эритроцитов и изменении их качественного состава. Любая анемия приводит к снижению дыхательной функции крови и развитию кислородного голодания тканей, что чаще всего выражается такими симптомами как бледность кожных покровов, повышенная утомляемость, слабость, головные боли, головокружение, учащенное сердцебиение, одышка и другими.
При рутинном исследовании мазка периферической крови морфолог указывает на отклонение размера эритроцитов в меньшую сторону (микроцитоз) или в большую сторону (макроцитоз), но такая оценка, если она производится без специальных приспособлений — микрометров, — не может быть свободна от субъективизма. Достоинством автоматического анализа крови является стандартизация показателя — средний корпускулярный объем (СКО), измеряемый в фемтолитрах (fl, фл). СКО измеряют с помощью автоматизированного счетчика прямым способом. Нормальное значение среднего корпускулярного объема представляет 80—90 фл (нормоцитоз). Повышение СКО выше 95 фл — макроцитоз. Снижение СКО менее 80—75 фл — микроцитоз. Недостатком является относительно дорогое и сложное оборудование, требующее соответствующего сервисного обслуживания.
Замена цветового показателя на показатель СКО не нарушает привычной классификации анемий, построенной на основании цветового показателя.
Определение анемии
В зависимости от пола и возраста норма показателя содержания гемоглобина в литре крови может отличаться.
| Возраст и пол | Порог Hb(Г/Л) | Порог Hb(Г/%) |
| Дети (3 мес.-5 лет) | 110 | 11,0 |
| Дети (5-12 лет) | 115 | 11,5 |
| Дети (12-15 лет) | 120 | 12,0 |
| Мужчины(>15 лет) | 130-160 | 13,0-16,0 |
| Женщины небеременные(>15 лет) | 120-140 | 12,0-14,0 |
| Женщины беременные | 110 | 11,0 |
Классификация анемий
Анемии подразделяют на группы по различным признакам. Классификация анемий в основном основывается на удобстве, возможности эффективного её применения в клинической практике.
По цветовому показателю
Цветовой показатель (ЦП) показывает степень насыщения эритроцита гемоглобином. В норме он равен 0,85 — 1,05. В зависимости от него различают такие анемии:
Гипохромная анемия
-
Гипохромные — ЦП < 0,85(по некоторым источникам ниже 0,8):-
железодефицитная анемия -
талассемия
-
Нормохромная анемия
-
Нормохромные — ЦП 0,85-1,05:-
гемолитические анемии (когда скорость разрушения эритроцитов превышает скорость их продукции) -
постгеморрагическая (как результат потери крови вследствие кровотечения или кровоизлияния) -
неопластические заболевания костного мозга -
апластические анемии -
внекостномозговые опухоли -
анемии вследствие снижения выработки эритропоэтина
-
Гиперхромная анемия
-
Гиперхромные — ЦП > 1,1:-
витамин B12-дефицитная анемия -
фолиеводефицитная анемия -
миелодиспластический синдром
-
По степени тяжести
В зависимости от выраженности снижения уровня гемоглобина выделяют три степени тяжести анемии:
-
Лёгкая — уровень гемоглобина ниже нормы, но выше 90 г/л; -
Средняя — гемоглобин в пределах 90-70 г/л; -
Тяжёлая — уровень гемоглобина менее 70 г/л.

По способности костного мозга к регенерации
Основным признаком такой регенерации является увеличение количества ретикулоцитов (молодых эритроцитов) в периферической крови. Норма — 0,5 %-2 %
-
Арегенераторная (к примеру, апластическая анемия) — характерно отсутствие ретикулоцитов. -
Гипорегенераторная (витамин B12-дефицитная анемия, железодефицитная анемия) — характерно количество ретикулоцитов ниже 0,5 %. -
Норморегенераторная или регенераторная (постгеморрагическая) — количество ретикулоцитов в норме (0,5 %-2 %) -
Гиперрегенераторная (гемолитические анемии) — количество ретикулоцитов более 2 %
Патогенетическая классификация
Основана на механизмах развития анемий как патологического процесса
-
Дисгемопоэтические анемии — анемии, связанные с нарушением кровообразования в красном костном мозге -
Анемии, связанные с острой или хронической кровопотерей -
Анемии, связанные с повышенным гемолизом
Регуляция кроветворения
Регуляция кроветворения осуществляется главным образом гуморальным путем. Причем для каждого из рядов кроветворения, видимо, этот путь является самостоятельным. В отношении эритроцитопоэза известно, что дифференцировка поэтиночувствительных клеток в эритробласты (с последующими их дифференцировками до зрелых эритроцитов) невозможна без эритропоэтина. Стимулятором для выработки эритропоэтина является падение напряжения кислорода в тканях. Для дифференцировки гранулоцитов в культуре необходимо присутствие колониестимулирующего фактора, который, как и эритропоэтин, относится к а2-глобулинам.
Кроме специфических гормонов типа эритропоэтина, на кроветворение действуют и другие гормоны, например андрогены. Они стимулируют эритроцитопоэз, мобилизуя эндогенный эритропоэтин. Медиаторы (адреналин, ацетилхолин) влияют на кроветворную систему, не только вызывая перераспределение форменных элементов в крови, но и путем прямого воздействия на стволовые клетки (у них обнаружены адрено- и холинорецепторы).
Мало разработан вопрос о нервной регуляции кроветворения, хотя обильная иннервация кроветворных тканей не может не иметь биологического значения. Нервное напряжение, эмоциональные перегрузки ведут к развитию кратковременного нейтрофильного лейкоцитоза без существенного омоложения состава лейкоцитов. Несколько повышает уровень лейкоцитов в крови прием пищи. Аналогичный эффект вызывается введением адреналина. В основе этой реакции лежит преимущественно мобилизация сосудистого гранулоцитарного резерва. При этом лейкоцитоз развивается в течение нескольких десятков минут. Лейкоцитоз с палочкоядерным сдвигом вызывается введением пирогенала и глюкокортикоидных стероидных гормонов, достигая максимума через 2—6 час, и обусловлен выходом гранулоцитов из костномозгового резерва. Содержание гранулоцитов в костномозговом резерве превышает их количество в кровяном русле в 30—50 раз.
Гуморальная регуляция кроветворения осуществляется преимущественно на уровне поэтиночувствительных клеток. В опытах с неравномерным облучением было показано, что восстановление кроветворных клеток в облученной конечности происходит независимо от состава крови и состояния необлученных участков костного мозга. Пересадка костного мозга под капсулу мышиной почки показала, что объем костного мозга, развивающегося из трансплантата, определяется количеством пересаженных стромальных клеток. Следовательно, они и определяют пределы размножения стволовых клеток, из которых затем развивается костный мозг в почке мыши-реципиента. Работами А. Я. Фриденштенна и др. (1968, 1970) показана специфичность стромальных клеток различных кроветворных органов: стромальные клетки селезенки определяют дифференцировку стволовых клеток в направлении лимфоцитопоэза, костномозговые стромальные клетки — в направлении миелопоэза. Вместе с тем, по-видимому, существуют мощные стимуляторы, включение которых происходит при необычных состояниях (например, резкая анемия), что приводит к развитию в селезенке очагов несвойственного ей кроветворения с преимущественным размножением эритрокариоцитов. Чаще это наблюдается в детском возрасте. Такие очаги кроветворения, называемые экстрамедуллярными, содержат наряду с эритрокариоцитами небольшой процент других элементов костного мозга — миелоцитов, промиелоцитов, мегакариоцитов. При острой массивной или при длительной повышенной потере клеток кроветворение может идти по дополнительным путям в каждом из рядов. По-видимому, существуют возможности к появлению особых клеток-предшественниц 3-го ряда схемы кроветворения, которые и дают начало таким шунтовым путям кроветворения, обеспечивающим быструю продукцию большого количества клеток. Это хорошо прослежено при эритроцитопоэзе, но, вероятно, существует и в других рядах.
Включение стволовых клеток в дифференцировку является скорее всего случайным процессом, вероятность которого при стабильном кроветворении составляет примерно 50%. Регуляция числа стволовых клеток носит не общий, а локальный характер и обеспечивается механизмами, функционирующими в каждом конкретном участке кроветворного микроокружения. Значительно менее ясно, регулируется ли направление дифференцировки стволовых кроветворных клеток. На основании целого ряда экспериментальных данных высказываются предположения о том, что вероятность дифференцировки стволовых клеток в направлении эритроцитопоэза, гранулоцитопоэза и т. д. всегда постоянна и не зависит от внешних условии.
Фактов, свидетельствующих о существовании специализированной системы, регулирующей кроветворение, нет. Поддержание определенного количества зрелых клеток в крови осуществляется многоступенчатой передачей нейрогуморальных сигналов. Сигнал поступает к клеточному резерву или клеточному депо, из которого эритроциты мобилизуются очень быстро при острой кровопотере. Затем стимулируется продукция соответствующих клеток на уровне поэтиночувствительных элементов путем увеличения их численности сначала без дифференцировки («горизонтальные митозы»), а затем с дифференцировкой. В результате создается категория зрелых клеток.
| |
| Периоды кроветворения Кроветворение в антенатальном периоде впервые обнаруживается у 19-дневного эмбриона в кровяных островках желточного мешка, в стебле и хорионе. К 22-му дню первые кровяные клетки проникают в мезодермальную ткань эмбриона, в сердце, аорту, артерии. На 6-й неделе снижается активность кроветворения в желточном мешке. Полностью первый (мезобластический) период гемопоэза, преимущественно эритроцитопоэза, заканчивается к началу 4-го месяца жизни эмбриона. Примитивные кроветворные клетки желточного мешка накапливают гемоглобин и превращаются в примитивные эритробласты, названные П. Эрлихом мегалобластами. Второй (печеночный) период кроветворения начинается после 6 неделе и достигает максимума к 5-му мес. К. этого периода преимущественно эритроидное, хотя на 9-й неделе в печени уже созревают первые нейтрофилы. Печеночный период эритроцитопоэза характеризуется исчезновением мегалобластов; при этом эритрокариоциты имеют нормальные размеры. На 3-м мес. эмбриональной жизни в эритроцитопоэз включается селезенка, но у человека ее роль в пренатальном кроветворении ограничена. На 4—5-м мес. начинается третий (костномозговой) период кроветворения Миелоидный эритроцитопоэз плода — эритробластический и, как и лейкоцитопоэз, мало отличается от эритроцитопоэза взрослого. Общей закономерностью эмбрионального эритроцитопоэза является постепенное уменьшение размеров эритроцитов и увеличение их числа. Соответственно различным периодам кроветворения (мезобластическому, печеночному и костномозговому) существует три разных типа гемоглобина: эмбриональный, фетальный и гемоглобин взрослого. В основном переход от фетального гемоглобина к гемоглобину взрослого начинается на 3-й нед. жизни плода и заканчивается через 6 мес. после рождения. В первые дни у новорожденных наблюдается полиглобулия и нейтрофильный лейкоцитоз. Затем активность эритроцитопоэза снижается. Нормализуется он в возрасте 2—3 мес. Нейтрофилез первых дней жизни сменяется лимфоцитозом; только к 5 годам в лейкоцитарной формуле начинают преобладать нейтрофилы.
ТромбоцитопоэзСамой молодой клеткой тромбоцитопоэза (тромбопоэза) является мегакариобласт — одноядерная небольшая клетка с крупным бластным ядром, хроматинные нити которого толще и грубее, чем у эритробласта; в ядре могут быть видны 1—2 темно-синие нуклеолы. Цитоплазма беззернистая, темно-синего цвета, отростчатая, узким ободком окружает ядро. Промегакариоцит возникает в результате нескольких эндомитозов. Ядро полиморфное с грубым строением хроматина; цитоплазма темно-синяя, беззернистая. Зрелый мегакариоцит отличается от промегакариоцита большим ядром. Цитоплазма имеет сине-розовую окраску, содержит азурофильную красноватую зернистость. Внутри мегакариоцита формируются тромбоциты. В мазке можно видеть и распадающиеся мегакариоциты, окруженные кучками тромбоцитов. При тромбоцитолитических состояниях отшнуровка тромбоцитов может происходить и на стадии промегакариоцита, тромбоциты при этом лишены азурофильной субстанции, но они активно участвуют в гемостазе. ЭритропоэзВ эритроцитопоэзе (эритропоэзе) самой молодой клеткой является эритробласт (ее называют также проэритробластом), который имеет бластную структуру и обычно круглое ядро. Цитоплазма при окраске темно-синяя, располагается узким ободком, часто дает своеобразные выросты. В отношении клеток эритрокариоцитарного ряда нет единой номенклатуры. Одни называют их нормобластами, другие эритробластами. Поскольку для других рядов термин «бласт» применяется лишь для клеток—родоначальниц того или иного ростка (отсюда и название «бласт» — росток), все клетки, являющиеся потомством эритробласта, должны иметь в названии окончание «цит». Поэтому термин «нормобласты» был заменен на «нормоциты». За эритробластом появляется пронормоцит, который отличается от эритробласта более грубым строением ядра, хотя оно и сохраняет правильную структуру хроматиновых нитей. Диаметр ядра меньше, чем у эритробласта, ободок цитоплазмы шире, и становится видна перинуклеарная зона просветления. При изучении миелограммы пронормоцит легко спутать с эритробластом. В связи с трудностью разделения этих клеток некоторые авторы предлагают в практической гематологии их вообще не дифференцировать. Далее располагается базофильный нормоцит, у которого грубоглыбчатое ядро имеет колесовидную структуру, цитоплазма окрашена в темно-синий цвет. Следующий — полихроматофильный — нормоцит имеет еще более плотную структуру ядра; цитоплазма занимает большую часть клетки и имеет базофильную окраску за счет структур, содержащих РНК, и оксифильную за счет появления уже достаточного количества гемоглобина. Ортохромный, или оксифильный, нормоцит имеет маленькое плотное ядро (как вишневая косточка), оксифильную или с базофильным оттенком цитоплазму. В норме оксифильных нормоцитов сравнительно мало, т. к., выталкивая на этой стадии ядро, клетка превращается в эритроцит, но в «новорожденном» эритроците всегда сохраняются остатки базофилии за счет небольшого количества РНК, которая исчезает в течение первых суток. Такой эритроцит с остатками базофилии называется полихроматофильным эритроцитом. При применении специальной прижизненной окраски базофильное вещество выявляется в виде сеточки; тогда эту клетку называют ретикулоцитом. Зрелый эритроцит имеет форму двояковогнутого диска, поэтому в мазке крови он имеет центральное просветление. По мере старения форма эритроцита постепенно приближается к сферической. |