Файл: Годфруа Ж. Что такое психология в 2х т. Оглавление предисловие редактора перевода.doc
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 16.03.2024
Просмотров: 896
Скачиваний: 0
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.

рефрактерности,в течение которого мембрана реполяризуется и восстанавливает способность к проведению нового импульса*. Так за счет последовательных деполяризаций и происходит распространение этогопотенциала действия(т. е. нервного импульса) с постоянной скоростью, варьирующей в пределах от 0,5 до 120 метров в секунду в зависимости от типа волокна, его толщины и наличия или отсутствия у него миелиновой оболочки.
* Во время периода рефрактерности, длящегося около тысячной доли секунды, нервные импульсы по волокну проходить не могут. Поэтому за одну секунду нервное волокно способно провести не более 1000 импульсов.
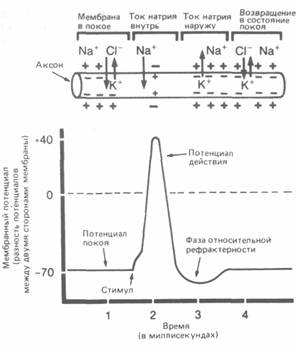
Рис. А.29. Потенциал действия. Развитие потенциала действия, сопровождающееся изменением электрического напряжения (от —70 до + 40 мВ), обусловлено восстановлением равновесия между положительными и отрицательными ионами по обе стороны мембраны, проницаемость которой на короткое время увеличивается.
Закон «всё или ничего».Поскольку каждому нервному волокну присущ определенный электрический потенциал, распространяющиеся по нему импульсы независимо от интенсивности или каких-либо других свойств внешнего стимула всегда имеют одни и те же характеристики. Это означает, что импульс в нейроне может возникнуть только в том случае, если его активация, вызванная стимуляцией рецептора или импульсом от другого нейрона, будет превосходить некий порог, ниже которого активация неэффективна; но, если порог достигнут, сразу же возникает «полномерный» импульс. Этот факт получил название закона «всё или ничего».
Синаптическая передача
Синапс.Синапсом называют область соединения между окончанием аксона одного нейрона и дендритами или телом другого. Каждый нейрон может образовать до 800-1000 синапсов с другими нервными клетками, а плотность этих контактов в сером веществе мозга составляет боле 600 млн. на 1 мм3 (рис. А.30)*.
*Это значит, что если за одну секунду отсчитывать по 1000 синапсов, то для их полного пересчета потребуется от 3 до 30 тысяч лет (Changeux, 1983, р. 75).
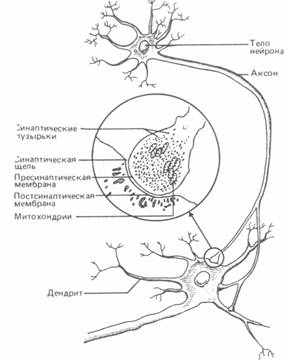
Рис. А.30. Синаптическое соединение нейронов (в середине - область синапса при большем увеличении). Терминальная бляшка пресинаптического нейрона содержит пузырьки с запасом нейромедиатора и митохондрии, доставляющие энергию, необходимую для передачи нервного сигнала.
Место перехода нервного импульса с одного нейрона на другой представляет собой, собственно, не точку контакта, а скорее узкий промежуток, называемыйсиноптической щелью.Речь идет о щели шириной от 20 до 50 нанометров (миллионных долей миллиметра), которая с одной стороны ограничена мембраной пресинаптической бляшки нейрона, передающего импульс, и с другой - постсинаптической мембраной дендрита или тела другого нейрона, принимающего нервный сигнал и затем передающего его дальше.
Нейромедиаторы.Именно в синапсах происходят процессы, в результате которых химические вещества, освобождаемые пресинаптической мембраной, передают нервный сигнал с одного нейрона на другой. Эти вещества, получившие название нейромедиаторов(или просто медиаторов),-своего рода «мозговые гормоны» (нейрогормоны) - накапливаются в пузырьках синаптических бляшек и освобождаются, когда по аксону сюда приходит нервный импульс.
После этого медиаторы диффундируют в синаптическую щель и присоединяются к специфическим рецепторным участкампостсинаптической мембраны, т. е. к таким участкам, к которым они «подходят, как ключ к замку». В результате этого проницаемость постсинаптической мембраны изменяется, и таким образом сигнал передается с одного нейрона на другой; медиаторы могут также и блокировать передачу нервных сигналов на уровне синапса, уменьшая возбудимость постси-наптического нейрона.
Выполнив свою функцию, медиаторы расщепляются или нейтрализуются ферментами либо всасываются обратно в пресинаптическое окончание, что приводит к восстановлению их запаса в пузырьках к моменту прихода следующего импульса (рис. А.31).
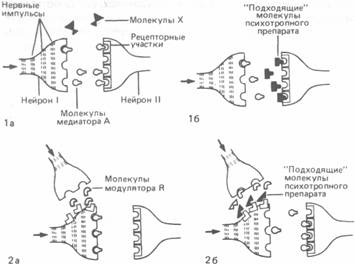
Рис. А.31. la. Медиатор А, молекулы которого освобождаются из концевой бляшки нейрона I, связывается специфическими рецепторами на дендритах нейрона II. Молекулы X, которые по своей конфигурации не подходят к этим рецепторам, занять их не могут и потому не вызывают каких-либо синаптических эффектов.
1б. Молекулы M (например, молекулы некоторых психотропных препаратов) сходны по своей конфигурации с молекулами нейромедиатора А и поэтому могут связываться с рецепторами для этого медиатора, таким образом мешая ему выполнять свои функции. Например, ЛСД мешает серотонину подавлять проведение сенсорных сигналов.
2а и 2б. Некоторые вещества, называемые нейромодуляторами, способны воздействовать на окончание аксона, облегчая или подавляя высвобождение нейромедиатора.

Возбуждающая или тормозная функция синапса зависит главным образом от типа выделяемого им медиатора и от действия последнего на постсинаптическую мембрану. Некоторые медиаторы всегда оказывают только возбуждающее действие, другие - только тормозное (ингибирующее), а третьи в одних отделах нервной системы играют роль активаторов, а в других-ингибиторов.
Функции главныхнейромедиаторов. В настоящее время известно несколько десятков этих нейрогормонов, но их функции изучены пока недостаточно. Сказанное, например, относится к ацетилхолину,который участвует в мышечном сокращении, вызывает замедление сердечного и дыхательного ритма и инактивируется ферментомацетилхолинэстеразой*. Не вполне изучены и функции таких веществ из группы моноаминов,как норадреналин, отвечающий за бодрствование мозговой коры и учащение сердечного ритма, дофамин,присутствующий в «центрах удовольствия» лимбической системы и некоторых ядрах ретикулярной формации, где он участвует в процессах избирательного внимания, или серотонин,который регулирует сон и определяет объем информации, циркулирующей в сенсорных путях. Частичная инактивация моноаминов происходит в результате их окисления ферментоммоноаминоксидазой.Этот процесс, обычно возвращающий активность мозга к нормальному уровню, в некоторых случаях может приводить к чрезмерному ее снижению, что в психологическом плане проявляется у человека в чувстве подавленности (депрессии).
* По-видимому, недостаток ацетилхолина в некоторых ядрах промежуточного мозга-одна из главных причин болезни Альцгеймера, а недостаток дофамина в скорлупе (одно из базальных ядер) может быть причиной болезни Паркиисона.
Гамма-аминомасляная кислота (ГАМК)представляет собой нейро-медиатор, выполняющий примерно ту же физиологическую функцию, что и моноаминоксидаза. Ее действие состоит главным образом в снижении возбудимости мозговых нейронов по отношению к нервным импульсам.
Наряду с нейромедиаторами существует группа так называемыхнейромодуляторов,которые в основном участвуют в регуляции нервного ответа, взаимодействуя с медиаторами и видоизменяя их эффекты. В качестве примера можно назвать вещество Ри брадикинин,участвующие в передаче болевых йпгналов. Освобождение этих веществ в синапсах спинного мозга, однако, может быть подавлено секрецией эндорфиновиэнкефалина,которая таким образом приводит к уменьшению потока болевых нервных импульсов (рис. А.31, 2а). Функции модуляторов выполняют и такие вещества, как
факторS,играющий, по-видимому, важную роль в процессах сна, холецистокинин,ответственный за чувство сытости, ангиотензин,регулирующий жажду, и другие агенты.
Нейромедиаторы и действие психотропных веществ.В настоящее время известно, что различные психотропные препаратыдействуют на уровне синапсов и тех процессов, в которых участвуют нейромедиаторы и нейромодуляторы.
Молекулы этих препаратов по своей структуре сходны с молекулами определенных медиаторов, что и позволяет им «обманывать» различные механизмы синаптической передачи. Таким образом они нарушают действие истинных нейромедиаторов, либо занимая их место на рецепторных участках, либо мешая им всасываться обратно в пресинаптические окончания или подвергаться разрушению специфическими ферментами (рис. А.31, 26).
Установлено, например, что ЛСД, занимая серотониновые рецепторные участки, мешает серотонину затормаживать приток сенсорных сигналов. Таким образом ЛСД открывает доступ к сознанию для самых разнообразных стимулов, непрерывно атакующих органы чувств.
Кокаинусиливает эффекты дофамина, занимая его место в рецепторных участках. Подобным же образом действуют морфини другие опиаты, мгновенный эффект которых объясняется тем, что они быстро успевают занять рецепторные участки для эндорфинов*.
* Несчастные случаи, связанные с передозировкой наркотиков, объясняются тем, что связывание чрезмерного количества, например, героина зндорфиновыми рецепторами в нервных центрах продолговатого мозга приводит к резкому угнетению дыхания, а иногда и к полной его остановке (Besson, 1988, Science et Vie, Hors serie, n° 162).
Действие амфетаминовобусловлено тем, что они подавляют обратное поглощение норадреналина пресинаптическими окончаниями. В результате накопление избыточного количества нейрогормона в синаптической щели приводит к чрезмерной степени бодрствования мозговой коры.
Принято считать, что эффекты так называемых транквилизаторов(например, валиума) объясняются главным образом их облегчающим влиянием на действие ГАМК в лимбической системе, что приводит к усилению тормозных эффектов этого медиатора. Наоборот, как антидепрессантыдействуют главным образом ферменты, инактивирую-шие ГАМК, или такие препараты, как, например, ингибиторы моноаминоксидазы,введение которых увеличивает количества моноаминов в синапсах.
Смерть от некоторых
отравляющих газовнаступает вследствие удушья. Такое действие этих газов связано с тем, что их молекулы блокируют секрецию фермента, разрушающего ацетилхолин. Между тем ацетилхолин вызывает сокращение мышц и замедление сердечного и дыхательного ритма. Поэтому его накопление в синаптических пространствах приводит к угнетению, а затем и полной блокаде сердечной и дыхательной функций и одновременному повышению тонуса всей мускулатуры.
Изучение нейромедиаторов еще только начинается, и можно ожидать, что в скором времени будут открыты сотни, а может быть и тысячи этих веществ, многообразные функции которых определяют их первостепенную роль в регуляции поведения.
До недавнего времени единственным методом, позволяющим регистрировать электрическую активность мозга с помощью электродов, размещенных в разных участках черепной коробки, была электроэнцефалография (см. документ 4.1). Но записи, которые получают этим методом, с трудом поддаются расшифровке, и поэтому чаще всего электроэнцефалография дает лишь грубое представление об активности популяции нейронов, расположенных под электродом.
Недавно, однако, появилось другое устройство для регистрации нервной активности. Речь идет о так называемом сканере, позволяющем составлять довольно точные карты нервной активности в различных областях головного мозга.
Это устройство осуществляет томографическое сканирование головного мозга с помощью позитронной эмиссии (откуда и другое название сканера - позитронно-эмиссионный томограф). В основе метода лежит то обстоятельство, что для работы мозга используется главным образом глюкоза: чем выше активность данного участка, тем больше глюкозы ему требуется для поддержания работы.
Первый из такого рода методов заключается в выявлении активных зон мозга после инъекции в кровь радиоактивных изотопов (например, фтора-18 или углерода-11), способных испускать положительно заряженные частицы, называемые позитронами.Столкновение позитронов с отрицательно заряженными электронами в нейронах сопровождается «взрывом», в результате которого образуются два разлетающихся в противоположных направлениях фотона (см. документ 5.2). Эти кванты света, число которых должно быть больше в усиленно снабжаемых кровью активных участках, улавливались затем камерой с фоточувствительными элементами, производившей таким образом послойный анализ головного мозга. После определения компьютером точки возникновения каждого «взрыва» информация-точка за точкой - выводилась на телевизионный экран с изображением последовательных срезов мозга (рис. А.32).
* Во время периода рефрактерности, длящегося около тысячной доли секунды, нервные импульсы по волокну проходить не могут. Поэтому за одну секунду нервное волокно способно провести не более 1000 импульсов.
Рис. А.29. Потенциал действия. Развитие потенциала действия, сопровождающееся изменением электрического напряжения (от —70 до + 40 мВ), обусловлено восстановлением равновесия между положительными и отрицательными ионами по обе стороны мембраны, проницаемость которой на короткое время увеличивается.
Закон «всё или ничего».Поскольку каждому нервному волокну присущ определенный электрический потенциал, распространяющиеся по нему импульсы независимо от интенсивности или каких-либо других свойств внешнего стимула всегда имеют одни и те же характеристики. Это означает, что импульс в нейроне может возникнуть только в том случае, если его активация, вызванная стимуляцией рецептора или импульсом от другого нейрона, будет превосходить некий порог, ниже которого активация неэффективна; но, если порог достигнут, сразу же возникает «полномерный» импульс. Этот факт получил название закона «всё или ничего».
Синаптическая передача
Синапс.Синапсом называют область соединения между окончанием аксона одного нейрона и дендритами или телом другого. Каждый нейрон может образовать до 800-1000 синапсов с другими нервными клетками, а плотность этих контактов в сером веществе мозга составляет боле 600 млн. на 1 мм3 (рис. А.30)*.
*Это значит, что если за одну секунду отсчитывать по 1000 синапсов, то для их полного пересчета потребуется от 3 до 30 тысяч лет (Changeux, 1983, р. 75).
Рис. А.30. Синаптическое соединение нейронов (в середине - область синапса при большем увеличении). Терминальная бляшка пресинаптического нейрона содержит пузырьки с запасом нейромедиатора и митохондрии, доставляющие энергию, необходимую для передачи нервного сигнала.
Место перехода нервного импульса с одного нейрона на другой представляет собой, собственно, не точку контакта, а скорее узкий промежуток, называемыйсиноптической щелью.Речь идет о щели шириной от 20 до 50 нанометров (миллионных долей миллиметра), которая с одной стороны ограничена мембраной пресинаптической бляшки нейрона, передающего импульс, и с другой - постсинаптической мембраной дендрита или тела другого нейрона, принимающего нервный сигнал и затем передающего его дальше.
Нейромедиаторы.Именно в синапсах происходят процессы, в результате которых химические вещества, освобождаемые пресинаптической мембраной, передают нервный сигнал с одного нейрона на другой. Эти вещества, получившие название нейромедиаторов(или просто медиаторов),-своего рода «мозговые гормоны» (нейрогормоны) - накапливаются в пузырьках синаптических бляшек и освобождаются, когда по аксону сюда приходит нервный импульс.
После этого медиаторы диффундируют в синаптическую щель и присоединяются к специфическим рецепторным участкампостсинаптической мембраны, т. е. к таким участкам, к которым они «подходят, как ключ к замку». В результате этого проницаемость постсинаптической мембраны изменяется, и таким образом сигнал передается с одного нейрона на другой; медиаторы могут также и блокировать передачу нервных сигналов на уровне синапса, уменьшая возбудимость постси-наптического нейрона.
Выполнив свою функцию, медиаторы расщепляются или нейтрализуются ферментами либо всасываются обратно в пресинаптическое окончание, что приводит к восстановлению их запаса в пузырьках к моменту прихода следующего импульса (рис. А.31).
Рис. А.31. la. Медиатор А, молекулы которого освобождаются из концевой бляшки нейрона I, связывается специфическими рецепторами на дендритах нейрона II. Молекулы X, которые по своей конфигурации не подходят к этим рецепторам, занять их не могут и потому не вызывают каких-либо синаптических эффектов.
1б. Молекулы M (например, молекулы некоторых психотропных препаратов) сходны по своей конфигурации с молекулами нейромедиатора А и поэтому могут связываться с рецепторами для этого медиатора, таким образом мешая ему выполнять свои функции. Например, ЛСД мешает серотонину подавлять проведение сенсорных сигналов.
2а и 2б. Некоторые вещества, называемые нейромодуляторами, способны воздействовать на окончание аксона, облегчая или подавляя высвобождение нейромедиатора.
Возбуждающая или тормозная функция синапса зависит главным образом от типа выделяемого им медиатора и от действия последнего на постсинаптическую мембрану. Некоторые медиаторы всегда оказывают только возбуждающее действие, другие - только тормозное (ингибирующее), а третьи в одних отделах нервной системы играют роль активаторов, а в других-ингибиторов.
Функции главныхнейромедиаторов. В настоящее время известно несколько десятков этих нейрогормонов, но их функции изучены пока недостаточно. Сказанное, например, относится к ацетилхолину,который участвует в мышечном сокращении, вызывает замедление сердечного и дыхательного ритма и инактивируется ферментомацетилхолинэстеразой*. Не вполне изучены и функции таких веществ из группы моноаминов,как норадреналин, отвечающий за бодрствование мозговой коры и учащение сердечного ритма, дофамин,присутствующий в «центрах удовольствия» лимбической системы и некоторых ядрах ретикулярной формации, где он участвует в процессах избирательного внимания, или серотонин,который регулирует сон и определяет объем информации, циркулирующей в сенсорных путях. Частичная инактивация моноаминов происходит в результате их окисления ферментоммоноаминоксидазой.Этот процесс, обычно возвращающий активность мозга к нормальному уровню, в некоторых случаях может приводить к чрезмерному ее снижению, что в психологическом плане проявляется у человека в чувстве подавленности (депрессии).
* По-видимому, недостаток ацетилхолина в некоторых ядрах промежуточного мозга-одна из главных причин болезни Альцгеймера, а недостаток дофамина в скорлупе (одно из базальных ядер) может быть причиной болезни Паркиисона.
Гамма-аминомасляная кислота (ГАМК)представляет собой нейро-медиатор, выполняющий примерно ту же физиологическую функцию, что и моноаминоксидаза. Ее действие состоит главным образом в снижении возбудимости мозговых нейронов по отношению к нервным импульсам.
Наряду с нейромедиаторами существует группа так называемыхнейромодуляторов,которые в основном участвуют в регуляции нервного ответа, взаимодействуя с медиаторами и видоизменяя их эффекты. В качестве примера можно назвать вещество Ри брадикинин,участвующие в передаче болевых йпгналов. Освобождение этих веществ в синапсах спинного мозга, однако, может быть подавлено секрецией эндорфиновиэнкефалина,которая таким образом приводит к уменьшению потока болевых нервных импульсов (рис. А.31, 2а). Функции модуляторов выполняют и такие вещества, как
факторS,играющий, по-видимому, важную роль в процессах сна, холецистокинин,ответственный за чувство сытости, ангиотензин,регулирующий жажду, и другие агенты.
Нейромедиаторы и действие психотропных веществ.В настоящее время известно, что различные психотропные препаратыдействуют на уровне синапсов и тех процессов, в которых участвуют нейромедиаторы и нейромодуляторы.
Молекулы этих препаратов по своей структуре сходны с молекулами определенных медиаторов, что и позволяет им «обманывать» различные механизмы синаптической передачи. Таким образом они нарушают действие истинных нейромедиаторов, либо занимая их место на рецепторных участках, либо мешая им всасываться обратно в пресинаптические окончания или подвергаться разрушению специфическими ферментами (рис. А.31, 26).
Установлено, например, что ЛСД, занимая серотониновые рецепторные участки, мешает серотонину затормаживать приток сенсорных сигналов. Таким образом ЛСД открывает доступ к сознанию для самых разнообразных стимулов, непрерывно атакующих органы чувств.
Кокаинусиливает эффекты дофамина, занимая его место в рецепторных участках. Подобным же образом действуют морфини другие опиаты, мгновенный эффект которых объясняется тем, что они быстро успевают занять рецепторные участки для эндорфинов*.
* Несчастные случаи, связанные с передозировкой наркотиков, объясняются тем, что связывание чрезмерного количества, например, героина зндорфиновыми рецепторами в нервных центрах продолговатого мозга приводит к резкому угнетению дыхания, а иногда и к полной его остановке (Besson, 1988, Science et Vie, Hors serie, n° 162).
Действие амфетаминовобусловлено тем, что они подавляют обратное поглощение норадреналина пресинаптическими окончаниями. В результате накопление избыточного количества нейрогормона в синаптической щели приводит к чрезмерной степени бодрствования мозговой коры.
Принято считать, что эффекты так называемых транквилизаторов(например, валиума) объясняются главным образом их облегчающим влиянием на действие ГАМК в лимбической системе, что приводит к усилению тормозных эффектов этого медиатора. Наоборот, как антидепрессантыдействуют главным образом ферменты, инактивирую-шие ГАМК, или такие препараты, как, например, ингибиторы моноаминоксидазы,введение которых увеличивает количества моноаминов в синапсах.
Смерть от некоторых
отравляющих газовнаступает вследствие удушья. Такое действие этих газов связано с тем, что их молекулы блокируют секрецию фермента, разрушающего ацетилхолин. Между тем ацетилхолин вызывает сокращение мышц и замедление сердечного и дыхательного ритма. Поэтому его накопление в синаптических пространствах приводит к угнетению, а затем и полной блокаде сердечной и дыхательной функций и одновременному повышению тонуса всей мускулатуры.
Изучение нейромедиаторов еще только начинается, и можно ожидать, что в скором времени будут открыты сотни, а может быть и тысячи этих веществ, многообразные функции которых определяют их первостепенную роль в регуляции поведения.
Дополнение А.4. Нервная активность и сканер
До недавнего времени единственным методом, позволяющим регистрировать электрическую активность мозга с помощью электродов, размещенных в разных участках черепной коробки, была электроэнцефалография (см. документ 4.1). Но записи, которые получают этим методом, с трудом поддаются расшифровке, и поэтому чаще всего электроэнцефалография дает лишь грубое представление об активности популяции нейронов, расположенных под электродом.
Недавно, однако, появилось другое устройство для регистрации нервной активности. Речь идет о так называемом сканере, позволяющем составлять довольно точные карты нервной активности в различных областях головного мозга.
Это устройство осуществляет томографическое сканирование головного мозга с помощью позитронной эмиссии (откуда и другое название сканера - позитронно-эмиссионный томограф). В основе метода лежит то обстоятельство, что для работы мозга используется главным образом глюкоза: чем выше активность данного участка, тем больше глюкозы ему требуется для поддержания работы.
Первый из такого рода методов заключается в выявлении активных зон мозга после инъекции в кровь радиоактивных изотопов (например, фтора-18 или углерода-11), способных испускать положительно заряженные частицы, называемые позитронами.Столкновение позитронов с отрицательно заряженными электронами в нейронах сопровождается «взрывом», в результате которого образуются два разлетающихся в противоположных направлениях фотона (см. документ 5.2). Эти кванты света, число которых должно быть больше в усиленно снабжаемых кровью активных участках, улавливались затем камерой с фоточувствительными элементами, производившей таким образом послойный анализ головного мозга. После определения компьютером точки возникновения каждого «взрыва» информация-точка за точкой - выводилась на телевизионный экран с изображением последовательных срезов мозга (рис. А.32).