Файл: Тема 1 клетка Вопрос Химический состав, организация плазмолеммы. Функции плазмолеммы.docx
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 17.03.2024
Просмотров: 386
Скачиваний: 0
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
Адгезия — способность клеток избирательно прикрепляться друг к другу или к компонентам внеклеточного матрикса.
Прикрепление клеток к молекулам адгезии внеклеточного матрикса реализуют точечные (фокальные) адгезионные контакты. В образовании контакта участвуют трансмембранные рецепторы — интегрины, объединяющие внеклеточные и внутриклеточные структуры. Фокальный контакт содержит также винкулин, a–актинин, талин и другие белки. Характер распределения макромолекул адгезии во внеклеточном матриксе (фибронектин, витронектин) определяет место окончательной локализации клетки в формирующейся ткани.
Клеточная адгезия важна для образования клеточных сообществ практически всех типов тканей. Молекулы адгезии специфичны для каждого типа ткани.
Вопрос 7. Десмосома и полудесмосома. Строение, значение, примеры локализации.
Адгезионные межклеточные контакты механически скрепляют клетки между собой. К адгезионным относятся промежуточный контакт (опоясывающая десмосома, zonula adherens), десмосома (macula adherens), полудесмосома. (прелюдия)
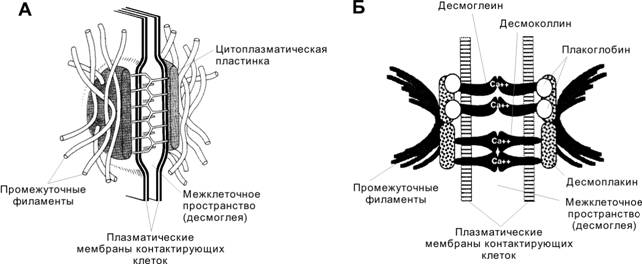
Десмосома — самый распространённый и наиболее сложно организованный тип межклеточных контактов. Десмосома содержит в себе две структуры: цитоплазматическую пластинку и десмоглию (внеклеточный мемжмебранный материал).
Цитоплазматичсеская пластинка связывает промежуточные филаменты клетки с плазматической мембраной.
Десмоглия связывается с цитоплазматической пластинкой и с десмоглией, которая контактирует с цитоплазматической пластинкой в соседней клетке.
Участки клеточных мембран, входящие в состав десмосомы, разделены слоем десмоглеи толщиной 20–30 нм. С внутренней стороны к плазматической мембране примыкает цитоплазматическая пластинка толщиной 10–40 нм с вплетёнными в неё промежуточными филаментами. В десмосомах всех клеточных типов присутствуют следующие белки: плакоглобин, десмоплакины, десмоглеины, десмоколлины.
Функция. Десмосомы поддерживают структурную целостность ткани, скрепляя клетки между собой. Десмосомы в комплексе с промежуточными филаментами придают ткани упругость и поддерживают в ней усилие натяжения.
Примеры. Десмосомы скрепляют клетки одного типа (кератиноциты, кардиомиоциты) и различных типов (тактильная клетка Меркеля–кератиноцит).
Полудесмосома обеспечивает прикрепление клетки к базальной мембране. Полудесмосома, как и десмосома, содержит цитоплазматическую пластинку с вплетёнными в неё промежуточными филаментами.
Вопрос 8. Адгезионный промежуточный контакт. Строение, примеры локализации.
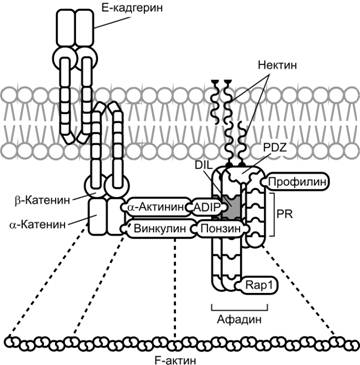
При промежуточном адгезионном контакте мембраны соседних клеток разделены промежутком шириной 10-20 нм, заполненным аморфным или фибриллярным материалом. Электроноплотная пластинка на цитоплазматической стороне клеточной мембраны в пределах контакта содержит белки плакоглобин, винкулин, a–актинин и радиксин. В пластинку вплетены концы актинсодержащих микрофиламентов. В образовании контакта участвуют трансмембранные белки адгезии из семейства кадгерина.
Цепь молекулярных взаимодействий выглядит следующим образом: актин (примембранные микрофиламенты) — a-актинин — винкулин — Е-кадгерин (трансмембранный гликопротеин адгезии). Е-кадгерин в присутствии Ca2+ прочно связывает мембраны соседних клеток. Цепь молекулярных взаимодействий в промежуточном контакте, начиная с актина микрофиламентов и до винкулина включительно, аналогична таковой в точечном (фокальном) адгезионном контакте. Если винкулин фокального контакта взаимодействует с трансмембранными белками-рецепторами (интегрины), то в промежуточном контакте он связывается с E-кадгерином.
В промежуточном контакте E-кадгерин связан с актиновым цитоскелетом через периферические мембранные белки, в том числе a-, b- и g–катенины, a–актинин и винкулин. b-Катенин взаимодействует с цитоплазматическим участком E-кадгерина и связывает E-кадгерин с a–катенином, который связан непосредственно с F–актином. a–Актинин и винкулин — также F–актин связывающие белки, прямо связанные с a–катенином. Связь E-кадгерина с актиновым цитоскелетом через эти мембранные белки усиливает межклеточную адгезионную активность E-кадгерина.
Другая молекула межклеточной адгезии, нектин, через белок афадин также связана с F–актином цитоскелета. Нектины (нектин-1, -2, -3 и -4) — Ca2+-независимые белки семейства иммуноглобулинов.
Афадин (белок AF6) связывает нектин с актиновым цитоскелетом. Необходим для совместной организации нектина и E-кадгерина в промежуточном контакте. Афадин связан с цитоплазматическим участком нектина и, через F–актинсвязывающий домен, с F–актином.
ADIP (Afadin DIL domain-Interacting Protein, SSX2-SSX2-interacting protein) — широко распространённый белок адгезии, участвующий в организации и присутствующий в промежуточных адгезионных контактах, связывает афадин и a–актинин. ADIP объединяет комплексы нектин–афадин и E-кадгерин–катенин с участием a–актинина. Кроме того, ADIP участвует в организации актинового цитоскелета в области промежуточного контакта через афадин и a–актинин.

Функция. Промежуточный контакт скрепляет не только мембраны соседних клеток, но и стабилизирует их цитоскелет, объединяя клетки с их содержимым в единую жёсткую систему.
Примеры: каёмчатый эпителий кишки (этот тип контактов известен как опоясывающая десмосома, т.к. контакт образует сплошной поясок вокруг клетки); секреторный эпителий (ацинозные клетки экзокринной части поджелудочной железы); вставочные диски в миокарде; эпендимные клетки ЦНС
Вопрос 9. Щелевой контакт. Структура, функция, примеры локализации.
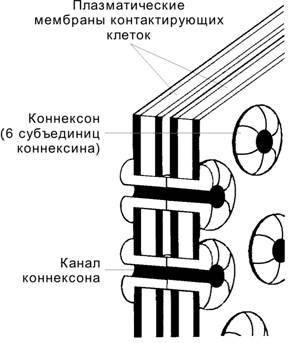
Щелевой контакт обеспечивает ионное и метаболическое сопряжение клеток. Плазматические мембраны клеток, образующих щелевой контакт, разделены щелью шириной 2–4 нм. Коннексон — трансмембранный белок цилиндрической конфигурации; состоит из 6 СЕ коннексина. Два коннексона соседних клеток соединяются в межмембранном пространстве и образуют канал между клетками.
Функция. Через щелевые контакты проходят низкомолекулярные вещества, регулирующие рост и развитие клеток. Для клеток нейроглии щелевой контакт имеет важное значение в регуляции уровня внутриклеточного Ca2+. Щелевые контакты обеспечивают распространение возбуждения — переход ионов между мышечными клетками миокарда и между ГМК.
Вопрос 10. Понятие о регенерации, физиологической и репаративной. Стволовые клетки, их значение.
Регенерация – восстановление утраченной или поврежденной дифференцированной структуры. Различают физиологическую и репаративную регенерацию.
Физиологическая регенерация – естественное обновление структуры. В ходе жизнедеятельности на смену гибнущим структурам приходят новые. В этом виде регенерации участвуют клетки всех обновляющихся популяции.
Репаративная регенерация – образование новых структур вместо поврежденных и на месте поврежденных. Возможна только в тканях из клеток обновляющихся популяций.
Стволовая клетка – недифференцированная клетка способная к самообновлению и дифференцировке в специализированные клетки.
Свойства:
-
Способна к самообновлению; -
Высокий пролиферативный потенциал; -
Способна к дифференцировке в клетки нескольких типов; -
Пластичность.
Характеристика стволовых клеток (СК):
-
Тотипотентность – клетка обладает дифференцироваться в любые клетки эмбриона и провизорных органов (зигота и бластомеры). -
Плюрипотентность – способность дифференцироваться в клетки, принадлежащие к трем зародышевым листкам (Клетки внутренней клеточной массы бластоцисты). -
Мультипотентность – способность дифференцироваться в клетки одного зародышевого листка (клетки лимфоидного и миелоидного кроветворных рядов). -
Унипотентность – способность дифференцироваться в клетки одного типа.
Вопрос 11. Классификация тканей и их общая характеристика.
Ткань – филогенетицчески сложившаяся система гистологических элементов, объединенных общей структурой, функцией и происхождением. Различают четыре основных тканевых типа: эпителиальная, ткани внутренней среды, мышечная и нервная ткани. В настоящее время принята классифиация тканей по фон Лейдигу.
Эпителиальная ткань.
Признаки:
-
Много клеток мало - межклеточного вещества. -
Наличие базальной мембраны. -
Наличие межклеточных контактов. -
Эпителий – бессосудистая ткань. -
Может происходить из всех зародышевых листков. -
Как правило хорошо регенерируют. -
Промежуточные филаменты – цитокератины. -
Наличие полярной дифференцировки. -
Пограничное положение. -
Пространственная организация.
Ткани внутренней среды. К ним относятся кровь, соединительная ткань и скелетная ткань.
Признаки:
-
Мало клеток много межклеточного вещества. -
Наличие волокон. -
Богатый видовой состав клеток. -
Способность большинства клеток к миграции. -
Способность большинства клеток к пролиферации. -
Как правило мезенхимное происхождение. -
Промежуточный филамент – белок вименти.
Мышечная ткань. Выделяют скелетную, сердечную и гладкую мышечную ткани.
Признаки.
-
Сократимость. -
Наличие акто–миозинового хемо-михонического преобразователя энергии. -
Мезодермальное происхождение. -
Промежуточный филамент – белок десмин.
Нервная ткань. Имеет невральное происхождение. Клеточные популяции: нейроны и клетки глии. Выполняет интегративную роль, координируя функции организма на всех уровнях его морфо-функциональной организации.
Тема 4: «ЭПИТЕЛИАЛЬНЫЕ ТКАНИ. ЭКЗОКРИННЫЕ ЖЕЛЕЗЫ»
Вопрос 1. Происхождение и функции эпителиальных тканей. Примеры.
Эпителиальная ткань образуется из всех трех зародышевых листков.
Транспорт газов (O2 и CO2) через эпителий альвеол лёгких; аминокислот и глюкозы при помощи специальных транспортных белков в эпителии кишки; IgA и других молекул на поверхность эпителиальных пластов.
Эндоцитоз, пиноцитоз. Эпителиальные клетки участвуют в пиноцитозе (например, эпителий почечных канальцев) и в опосредуемом рецепторами эндоцитозе (например, поглощение холестерина вместе с ЛНП или трансферрина большинством эпителиальных клеток).
Секреция. Экзоцитоз слизи, белков (гормонов, факторов роста, ферментов). Слизь вырабатывается специальными слизистыми клетками эпителия желудка и половых путей, бокаловидными клетками в эпителии кишки, трахеи и бронхов. Гормоны и факторы роста вырабатываются эндокринными клетками.
Барьерная. Разграничение сред путём образования надёжных барьеров из эпителиальных клеток, связанных плотными контактами (например, между эпителиальными клетками слизистой оболочки желудка и кишки).
Защита организма от повреждающего действия физических и химических факторов внешней среды.
Из эктодермы развивается эпителий эпидермиса, из мезодермы – эндокринные клетки надпочечников, из энтодермы - эпителий, выстилающий кишечную трубку, и железы, обеспечивающие пищеварение.
Вопрос 2. Основные черты организации пограничных эпителиев.
Основная роль пограничных эпителиев - отграничение организма от внешней среды и от вторичных полостей тела. Пограничные эпителии имеют пространственную организацию – пласт. Для клеток однослойного пласта характерна полярная дифференцировка, а многослойные пласты имеют значительные морфологические отличия между эпителиальными клетками разных слоёв.
Для эпителиальных пластов принята классификация, учитывающая количество слоёв клеток (одно- и многослойные), рядность однослойного эпителия (одно- и многорядные), форма клеток (для многослойных — поверхностного слоя), характер полярной дифференцировки.
Слойность.
Контакт всех клеток пласта с базальной мембраной определяет слойность эпителия. Если все клетки пласта связаны с базальной мембраной, эпителий — однослойный. Если это условие не выполняется, эпителий — многослойный. Эктодермальные эпителии — многослойные. Энтодермальные эпителии, как правило, — однослойные.
Рядность.
Рядность однослойных эпителиев отражает наличие (многорядный) или отсутствие (однорядный) в составе пласта клеток разной формы (в т.ч. разных типов клеток). По сути дела, этот