Файл: 7. Современное представление о центре кровообращения. Роль различных отделов центральнойнервной системы в регуляции кровообращения.docx
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 19.03.2024
Просмотров: 33
Скачиваний: 0
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
7. Современное представление о центре кровообращения. Роль различных отделов центральнойнервной системы в регуляции кровообращения


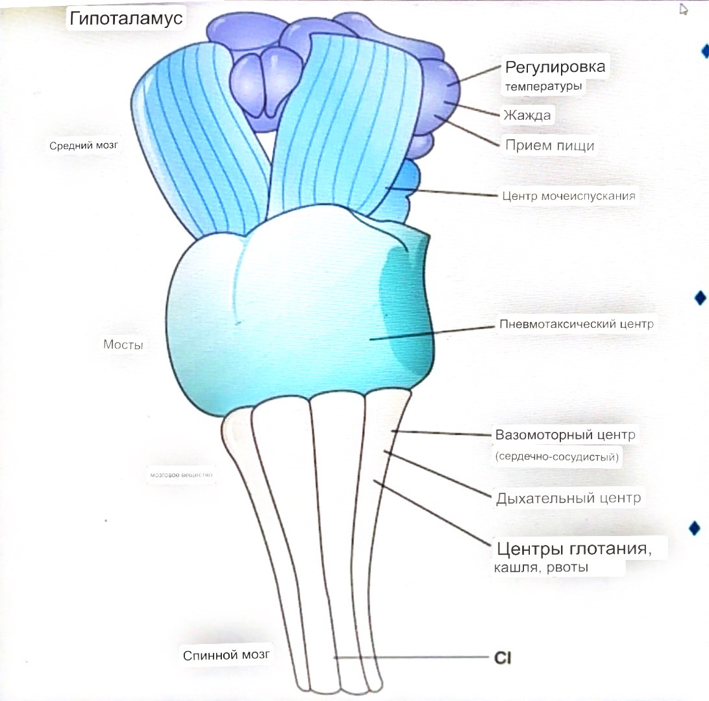
Сердечно-сосудистые центры ствола головного мозга
следует:
-
Сосудосуживающий центр (также называемый С1)
расположены в верхнем мозговом веществе и нижнем мосту.
Эфферентные нейроны этого сосудодвигательного центра входят в состав
симпатической нервной системы и синапсов в
спинной мозг, затем симпатические ганглии и
наконец, на органы-мишени, вызывая сужение сосудов в артериолах и венулах.
-
Кардиоускорительный центр.
Эфферентные нейроны из кардиоускорительного центра также являются частью
симпатическая нервная система и синапсы в
спинной мозг, симпатические ганглии и, наконец,
сердце. В сердце последствия этой деятельности
повышенная скорострельность узла SA (для увеличения частота сердечных сокращений), увеличение скорости проводимости через
атриовентрикулярный узел и повышенная сократимость.
-
Центр сердечного замедления.
Эфферентные волокна из центр сердечных замедлителей являются частью пара-
симпатическая нервная система: они путешествуют в блуждающий нерв и синапс на узле SA, чтобы уменьшить частота сердцебиения.
Сердечно-сосудистые центры ствола головного мозга расположены в ретикулярной формации продолговатого мозга и нижних одна треть моста. Эти центры функционируют в координированная мода, получение информации о крови давление от барорецепторов, а затем направление изменение активности симпатической и парасимпатической нервной системы, чтобы скорректировать кровяное давление, как нужный.
Как описано выше, артериальное давление определяется барорецепторами.
Также в каротидном синусе и дуге аорты. Афферентная
затем информация об артериальном давлении отправляется в продолговатый мозг через языкоглоточный (CN IX) и блуждающий (CN X) нервы. Эта информация интегрирована в
ядра солитарного тракта, которое затем направляет изменения
в деятельности ряда сердечно-сосудистых центров. Эти
сердечно-сосудистые центры тонически активны, а
ядро одиночного тракта просто направляет через центры увеличивается или уменьшается отток симпатических и парасимпатической нервной системы.
Парасимпатический отток является следствием
блуждающий нерв на узле SA, чтобы уменьшить частоту сердечных сокращений.
Симпатический отток состоит из четырех компонентов:
влияние на узел SA для увеличения частоты сердечных сокращений, эффект
на сердечную мышцу для повышения сократительной способности и инсульта
объем, воздействие на артериолы, вызывающее вазокон-стрикцию и увеличение TPR, а также воздействие на вены вызывают сужение вен и уменьшают ненапряженный объем.
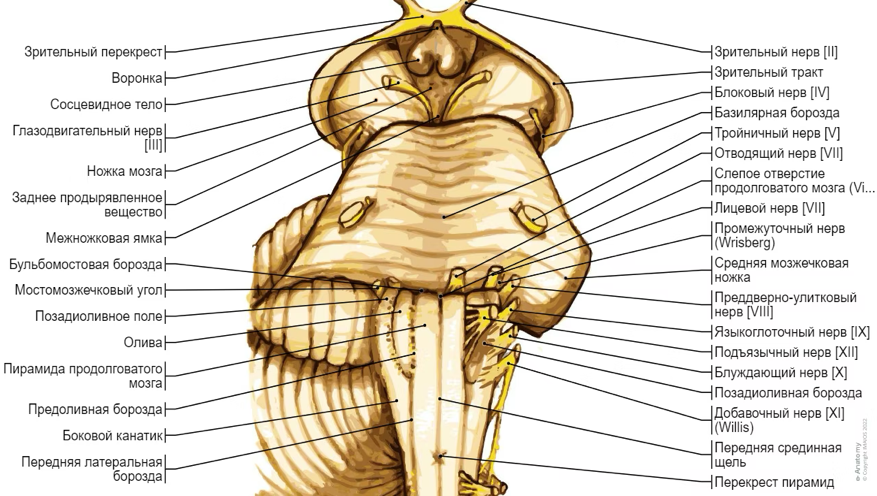
Все черепные нервы, кроме одного, отходят от
вентральная поверхность мозга; блоковый (IV) нерв берет начало
от дорсальной поверхности среднего мозга.
(
Лучше не получалось)
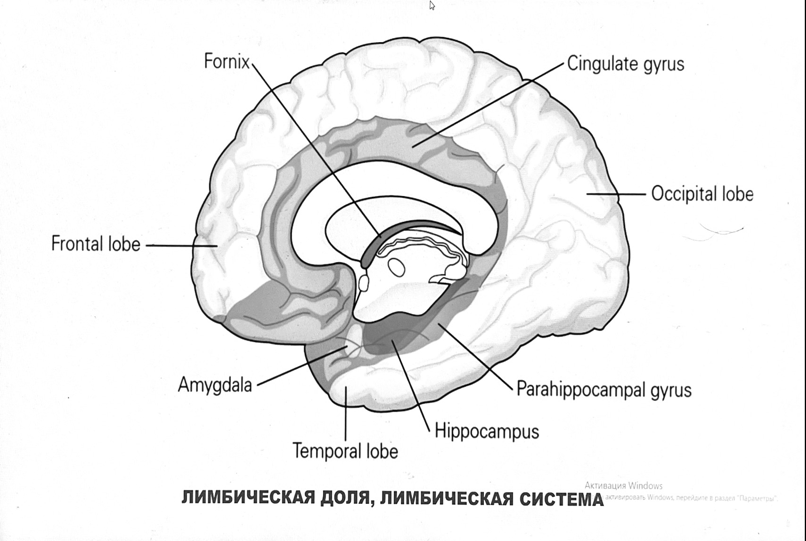
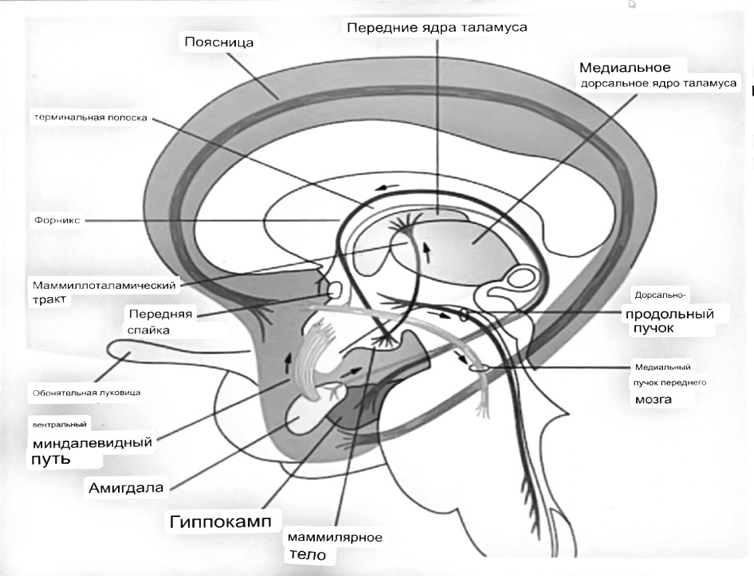
Выше-примитивная корковая ткань
Н
иже-г
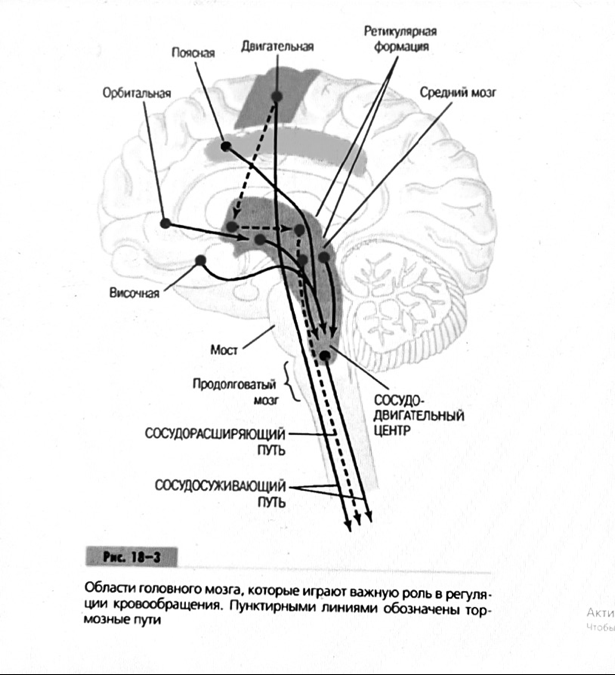
иппокамп,миндалевидное тело

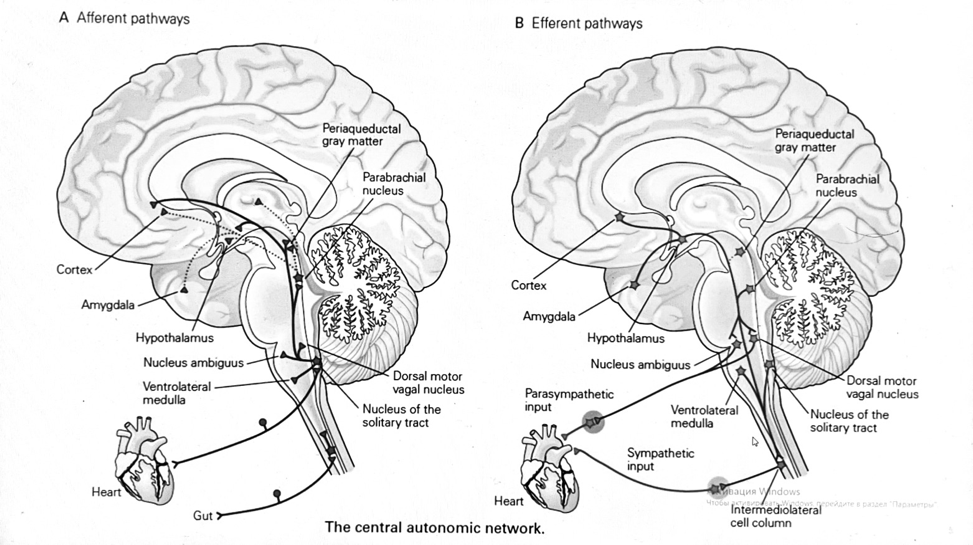
Вегетативная нервная система и артериальные барорецепторные рефлексы
синоатриальные и парасимпатические нервы иннервируют синоатриальный (СА) и атриовентрикулярный (АВ) узлы. Мио-
кардиальный отдел иннервируется симпатическими нервами. Артериальные и венозные сосуды на большей части тела
иннервируется симпатическими нервами, тогда как парасимпатическая нервная система иннервирует сосуды в
половых органов и желудочно-кишечного тракта. Вегетативная эфферентная активность регулируется барорецепторным рефлексом.
в ответ на изменение артериального давления, выявляемое барорецепторами каротидного синуса и дуги аорты.
проиллюстрирована реакция на изменение позы. АХ, ацетилхолин; АВ, атриовентрикулярный; ЦНС, центральная нервная
система; Е, адреналин; MAP, среднее артериальное давление; НЭ, норадреналин; СА, синоатриальный
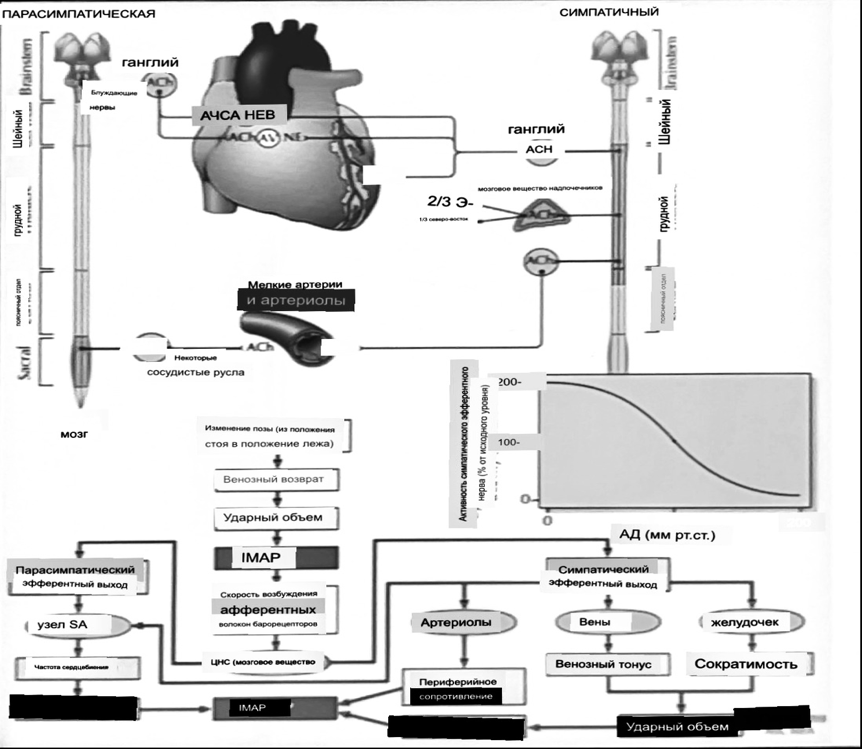
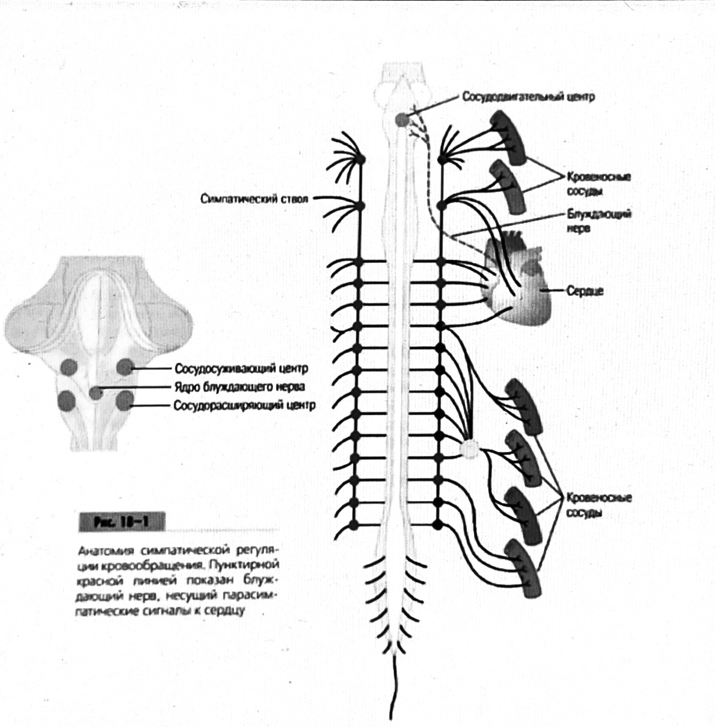
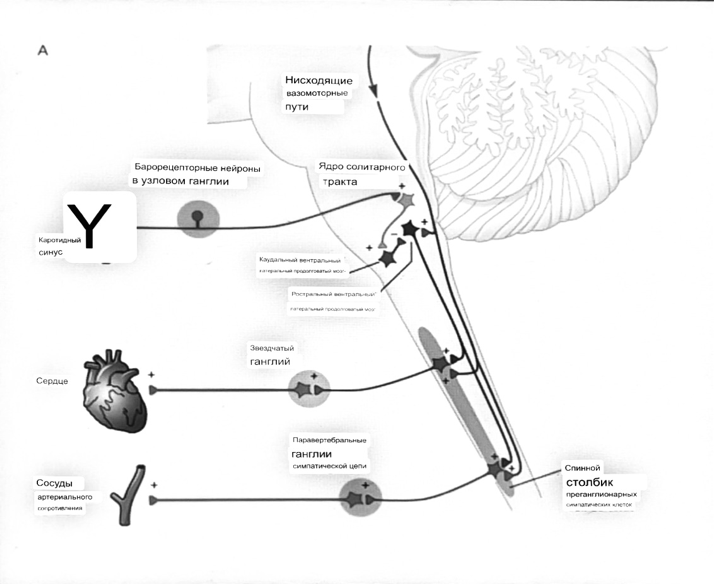
Общая схема центральной регуляции.

На рис.7.27 представлена общая схема организации центральных звеньев нервной регуляции кровообращения. Сердечно-сосудистый центр продолговатого мозга состоит из двух зон (А и Б), одна из которых определяет сужение сосудов (А'), ускорение и усиление сердцебиений (А"), тогда как другая — расширение сосудов (БI) и замедление частоты сокращений сердца (БII). Тонически активные нейроны зоны А непосредственно или через промежуточные нейроны возбуждает прегангли-онарные и ганглионарные симпатические нейроны, иннервирующие сердце и сосуды, и в то же время тормозят нейроны зоны Б. Последние активируются сигналами барорецепторов сердечно-сосудистой системы. Через короткие внутрицентральные пути нейроны
331
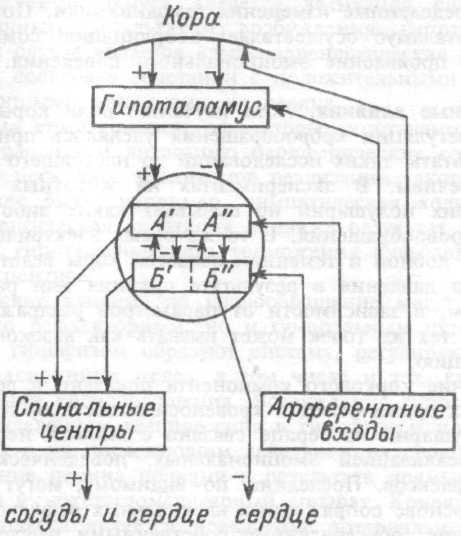
Рис.7.27. Схема организации центральных звеньев нервной регуляции сердечно-сосудистой системы. Объяснение в тексте.
зоны Б тормозят нейроны зоны А, а посредством нисходящих тормозных путей — и преганглионарные симпатические нейроны. Кроме того, через моторное ядро блуждающих нервов нейроны зоны Б влияют на ганглионарные парасимпатические нейроны сердца. В результате обе зоны (А и Б), получая основную сигнализацию от рецепторов сердечно-сосудистой системы, являются основой поддержания кровообращения, сочетательно подвергая сердце тоническому контролю тормозных вагусных и возбуждающих симпатических волокон, тогда как постоянный контроль сосудов обеспечивается только симпатическими вазоконстрикторными волокнами.
Афферентная нервная система является источником сигналов собственных и сопряженных рефлексов. Вход первых ограничен продолговатым мозгом, причем барорецепторы сердечно-сосудистой системы связаны с зоной Б, а хеморецепторы — с зоной А. С последней связаны также и некоторые афференты сопряженных рефлексов (от рецепторов кожи, мышц и внутренних органов), хотя организация этих связей известна лишь в общих чертах.
Поскольку сердечно- сосудистый центр продолговатого мозга регулирует основные параметры системной гемодинамики, даже будучи
отключенным от связей с вышележащими нервными структурами, он функционирует как истинный "центр", обладающий значительной независимостью. Следовательно, супрабульбарные структуры признаются несущественными ни для прямой тонической регуляции кровообращения, ни для осуществления сердечно-сосудистых рефлексов. В то же время при общих реакциях организма (поведенческих, эмоциональных, стрессорных и др.) кора мозга, гипоталамус и другие супрабульбарные структуры могут вмешиваться в деятельность сердечно-сосудистой системы, изменяя путем нисходящих (модулирующих) влияний возбудимость бульбарного центра.
Вазомоторные центры спинного мозга самостоятельного значения в регуляции кровообращения не имеют. Действующими элементами спинального уровня являются лишь симпатические преганглионар-ные нейроны, которые входят в состав эфферентного звена буль-барных рефлекторных дуг. Только в случае травм, приводящих к утрате связей продолговатого мозга со спинным, последний, спустя некоторое время, приобретает способность осуществлять вазомоторные рефлексы на импульсы спинномозговых афферентов.
8. Поддержание постоянства артериального давления в регуляции системного кровообращения.Срочные (рефлекторные) механизмы регуляции артериального давления. Рефлексы с основных рефлексогенных зон. Значение баро-, хемо-и волюмо-рецепторов. Собственные и сопряжённые рефлексы. Условно-рефлекторная регуляция кровообращения.
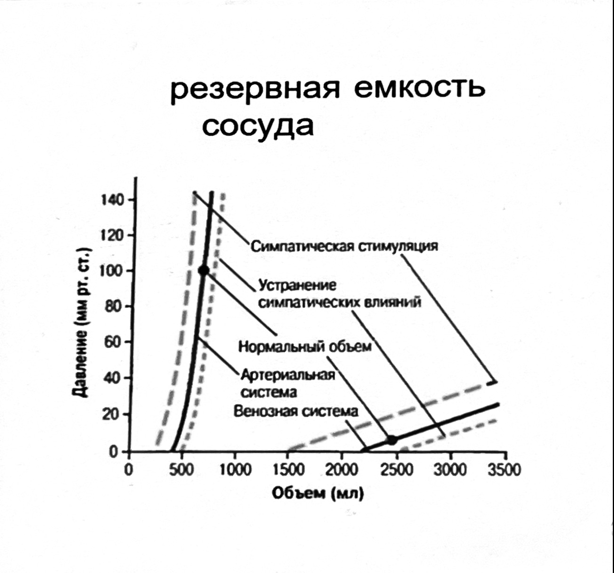
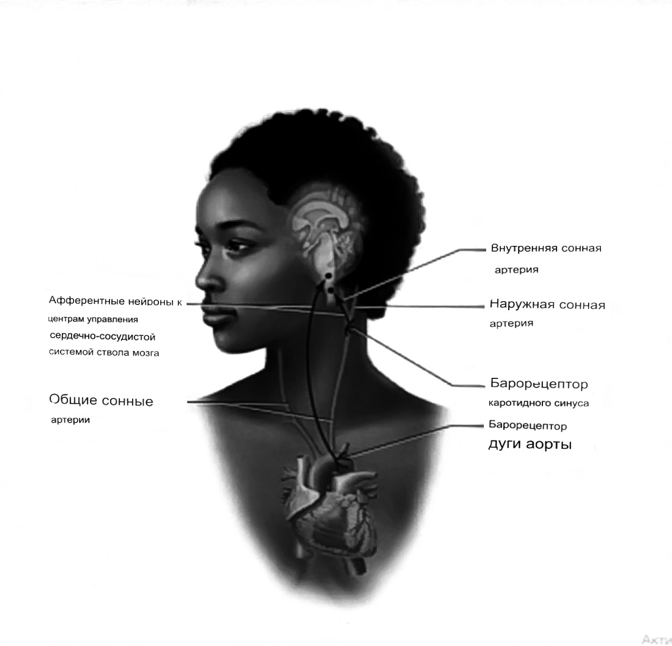
Значение баро-, хемо-и волюмо-рецепторов.


Возбуждение хеморецепторов аортальной и синокаротидной рефлексогенных зон возникает при уменьшении напряжения О2, увеличении напряжения СО2 и концентрации водородных ионов, т.е, при гипоксии, гиперкапнии и ацидозе. Импульсы от хеморецепторов поступают по тем же нервам, что и от барорецепторов, в продолговатый мозг, но непосредственно к нейронам прессорного отдела симпатического центра, возбуждение которого вызывает сужение сосудов, усиление и ускорение сердечных сокращений и как следствие — повышение АД. В результате кровь быстрее поступает к легким, углекислый газ обменивается на кислород.
Изменения деятельности сердечно-сосудистой системы способствуют устранению отклонений от нормы газового состава крови. Однако эффект невелик, так как увеличение АД осуществляется главным образом за счет сужения сосудов и лишь частично — в результате стимуляции деятельности сердца. Такой же эффект наблюдается и при нарушении кровоснабжения мозга любого происхождения (недостаточности деятельности сердца, нарушении мозгового кровообращения). Если степень нарушения кровоснабжения мозга выражена сильно, АД можети повыситься до 200 мм рт.ст. и больше.
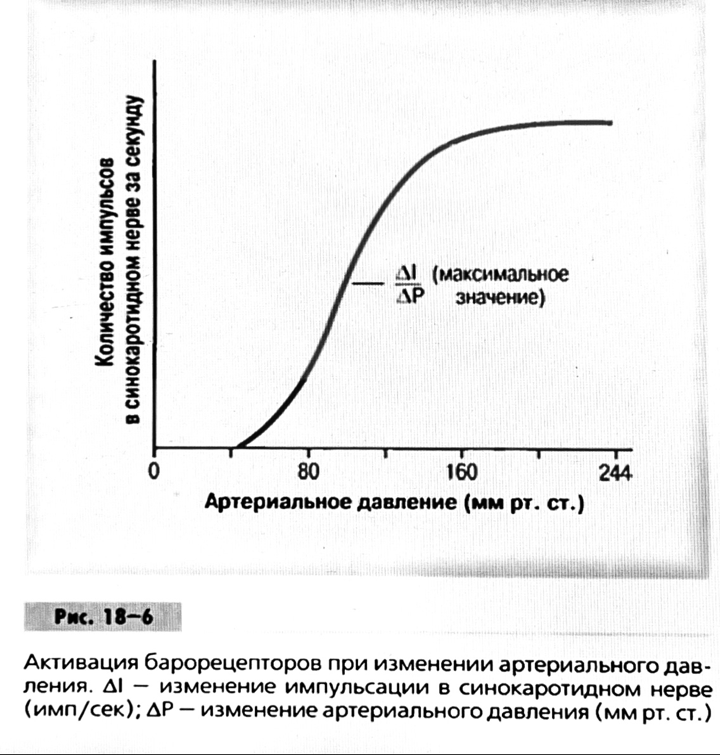
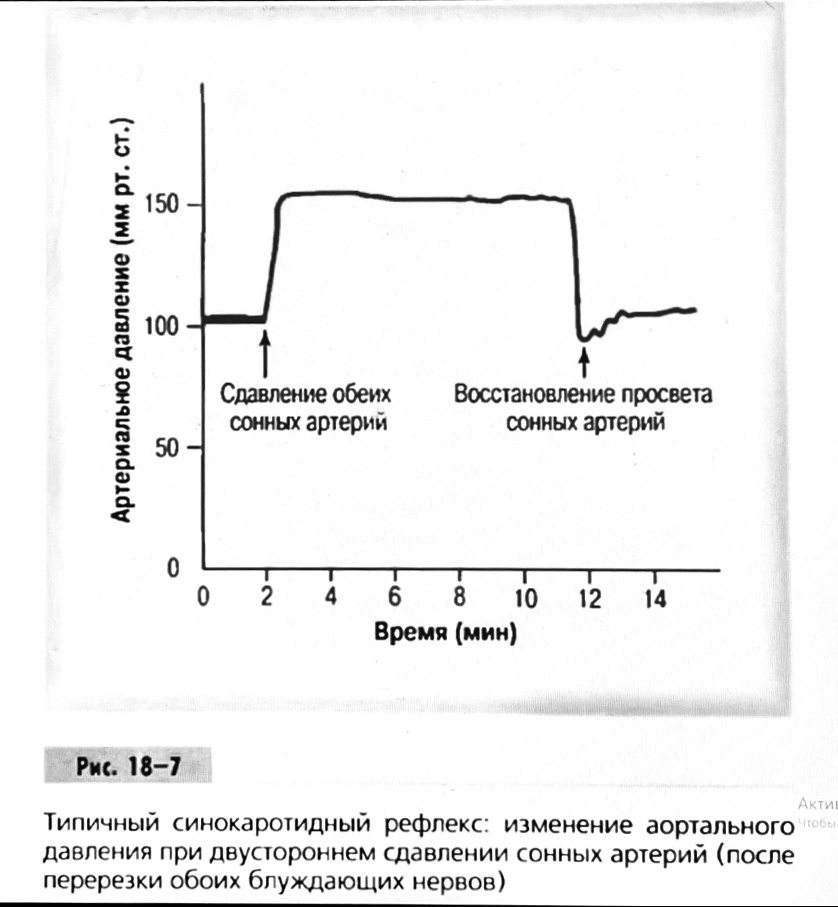

Барорецептор.
частота стрельбы меняется с изменением
артериальное давление. (a) Влияние изменения среднего
артериальное давление (MAP) в начале действия
потенциалы афферентными нейронами каротидной
синус. Этот эксперимент проводится путем накачивания
кровь непульсирующим образом через
изолированный каротидный синус, чтобы можно было установить
давление внутри него при любом желаемом значении.
(b) Активация потенциала действия барорецептора
частота колеблется в зависимости от давления. Увеличивать
при пульсовом давлении увеличивается общее действие
потенциальная частота даже при нормальном АД.
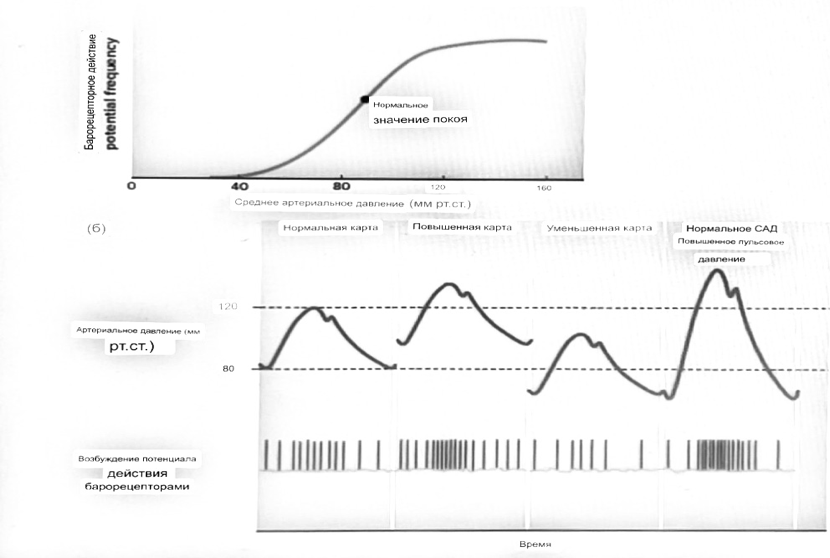
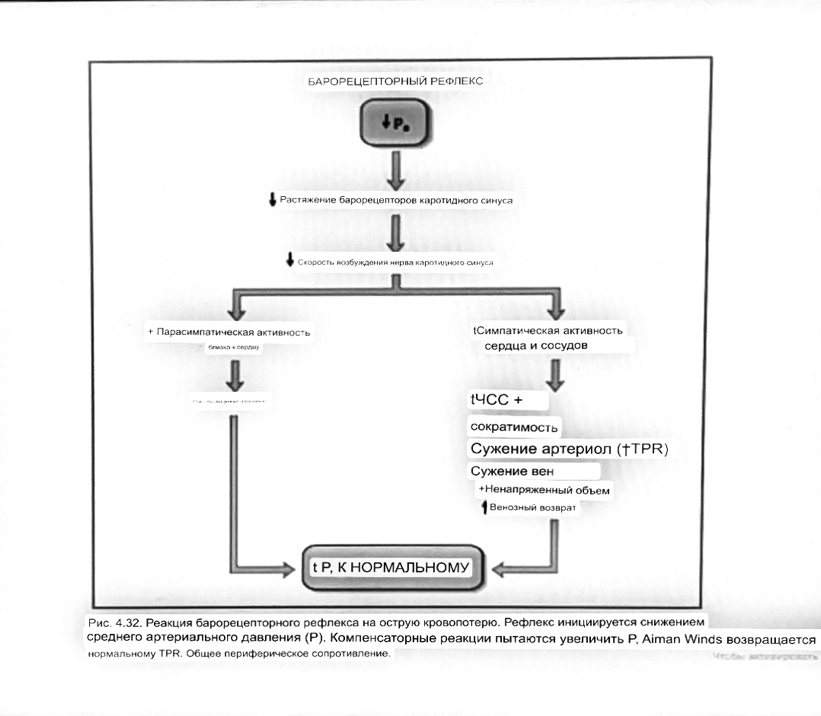

Барорецепторный рефлекс ведет себя как отрицательный
обратная связь с усилением.
A. Артериальное кровяное давление воспринимается барорецепторами, типом
рецепторный нейрон растяжения в каротидном синусе у основания
мозг. После интеграции в медулию эта информация про-
обеспечивает контроль сердечно-сосудистой системы с отрицательной обратной связью.
Симпатический компонент схемы включает в себя выходы
которые стимулируют насосную способность сердца (сердечный выброс)
за счет увеличения частоты сердечных сокращений и силы сокращений. В
кроме того, симпатическая стимуляция вызывает сокращение артерий.
что повышает гидравлическое сопротивление току крови. Вместе
последствия увеличения сердечного выброса и увеличения сосудистого
резистентность к повышению среднего артериального давления. Важно,
тормозные проекции от каудального отдела к ростральному вентральному
латеральный продолговатый мозг создает отрицательную обратную связь, так что увеличение
артериальное давление угнетает симпатическую активность, в то время как
снижение повышает симпатическую активность. Хотя опущено для
простота, парасимпатические нейроны в кардиальном узле также
способствуют рефлексу, создавая тормозной сердечный вход
который функционально антагонистичен симпатическому пути
(см. Рисунок 49-98). Во время барорецепторных рефлексов парасимпа-
Поэтому активность в сердце увеличивается за счет гипер-
напряжение и снижается при гипотонии,