ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 03.02.2024
Просмотров: 1068
Скачиваний: 0

СОДЕРЖАНИЕ
ПОПУЛЯЦИОННО-ВИДОВОЙ УРОВЕНЬ ОРГАНИЗАЦИИ ЖИЗНИ
ВИДООБРАЗОВАНИЕ В ПРИРОДЕ. ЭЛЕМЕНТАРНЫЕ ЭВОЛЮЦИОННЫЕ ФАКТОРЫ
11.5. ГЕНЕТИКО-АВТОМАТИЧЕСКИЕ ПРОЦЕССЫ (ДРЕЙФ ГЕНОВ)
11.7. НАСЛЕДСТВЕННЫЙ ПОЛИМОРФИЗМ ПРИРОДНЫХ ПОПУЛЯЦИЙ.
11.9. ПРОИСХОЖДЕНИЕ БИОЛОГИЧЕСКОЙ ЦЕЛЕСООБРАЗНОСТИ
ДЕЙСТВИЕ ЭЛЕМЕНТАРНЫХ ЭВОЛЮЦИОННЫХ ФАКТОРОВ
12.1. ПОПУЛЯЦИЯ ЛЮДЕЙ. ДЕМ, ИЗОЛЯТ
12.2. ВЛИЯНИЕ ЭЛЕМЕНТАРНЫХ ЭВОЛЮЦИОННЫХ ФАКТОРОВ НА ГЕНОФОНДЫ ЧЕЛОВЕЧЕСКИХ ПОПУЛЯЦИЙ
12.3. ГЕНЕТИЧЕСКОЕ РАЗНООБРАЗИЕ
13.1. ЭВОЛЮЦИЯ ГРУПП ОРГАНИЗМОВ
13.2. СООТНОШЕНИЕ ОНТО- И ФИЛОГЕНЕЗА
13.4. Организм как целое в историческом
Соотносительные преобразования органов
13.5. СОВРЕМЕННАЯ СИСТЕМА ОРГАНИЧЕСКОГО МИРА
ФИЛОГЕНЕЗ СИСТЕМ ОРГАНОВ ХОРДОВЫХ
14.2. ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
И ДАЛЬНЕЙШАЯ ЭВОЛЮЦИЯ ЧЕЛОВЕКА
15.3. ХАРАКТЕРИСТИКА ОСНОВНЫХ ЭТАПОВ АНТРОПОГЕНЕЗА
15.4. ВНУТРИВИДОВАЯ ДИФФЕРЕНЦИАЦИЯ ЧЕЛОВЕЧЕСТВА
БИОГЕОЦЕНОТИЧЕСКИЙ УРОВЕНЬ ОРГАНИЗАЦИИ ЖИЗНИ
16.1. БИОГЕОЦЕНОЗ - ЭЛЕМЕНТАРНАЯ ЕДИНИЦА БИОГЕОЦЕНОТИЧЕСКОГО УРОВНЯ ОРГАНИЗАЦИИ ЖИЗНИ
17.2. ЧЕЛОВЕК КАК ОБЪЕКТ ДЕЙСТВИЯ ЭКОЛОГИЧЕСКИХ ФАКТОРОВ.
АДАПТАЦИЯ ЧЕЛОВЕКА К СРЕДЕ ОБИТАНИЯ
17.4. РОЛЬ АНТРОПОГЕННЫХ ФАКТОРОВ
В ЭВОЛЮЦИИ ВИДОВ И БИОГЕОЦЕНОЗОВ
МЕДИЦИНСКАЯ ПАРАЗИТОЛОГИЯ. ОБЩИЕ ВОПРОСЫ
18.2. ФОРМЫ МЕЖВИДОВЫХ БИОТИЧЕСКИХ СВЯЗЕЙ В БИОЦЕНОЗАХ
18.3. КЛАССИФИКАЦИЯ ПАРАЗИТИЗМА
18.4. РАСПРОСТРАНЕННОСТЬ ПАРАЗИТИЗМА
18.5. ПРОИСХОЖДЕНИЕ ПАРАЗИТИЗМА
18.6. АДАПТАЦИИ К ПАРАЗИТИЧЕСКОМУ ОБРАЗУ ЖИЗНИ. ОСНОВНЫЕ ТЕНДЕНЦИИ
18.8. ФАКТОРЫ ВОСПРИИМЧИВОСТИ ХОЗЯИНА К ПАРАЗИТУ
18.9. ДЕЙСТВИЕ ХОЗЯИНА НА ПАРАЗИТА
18.10. СОПРОТИВЛЕНИЕ ПАРАЗИТОВ РЕАКЦИЯМ ИММУНИТЕТА ХОЗЯИНА
18.11. ВЗАИМООТНОШЕНИЯ В СИСТЕМЕ ПАРАЗИТ - ХОЗЯИН НА УРОВНЕ ПОПУЛЯЦИЙ
18.12. СПЕЦИФИЧНОСТЬ ПАРАЗИТОВ ПО ОТНОШЕНИЮ К ХОЗЯИНУ
18.13. ПРИРОДНО-ОЧАГОВЫЕ ЗАБОЛЕВАНИЯ
19.2. Простейшие, обитающие в полостных органах, сообщающихся с внешней средой
19.3. Простейшие, обитающие в тканях
19.4. ПРОСТЕЙШИЕ — ФАКУЛЬТАТИВНЫЕ ПАРАЗИТЫ ЧЕЛОВЕКА
20.1. ТИП ПЛОСКИЕ ЧЕРВИ PLATHELMINTHES
20.2. ТИП КРУГЛЫЕ ЧЕРВИ NEMATHELMINTHES
21.1. КЛАСС ПАУКООБРАЗНЫЕ ARACHNOIDEA
ЭВОЛЮЦИЯ ПАРАЗИТОВ И ПАРАЗИТИЗМА ПОД ДЕЙСТВИЕМ АНТРОПОГЕННЫХ ФАКТОРОВ
23.1. ПРОИСХОЖДЕНИЕ ЯДОВИТОСТИ
23.2. ЧЕЛОВЕК И ЯДОВИТЫЕ ЖИВОТНЫЕ
24.1. СОВРЕМЕННЫЕ КОНЦЕПЦИИ БИОСФЕРЫ
24.2. СТРУКТУРА И ФУНКЦИИ БИОСФЕРЫ
25.2. ПУТИ ВОЗДЕЙСТВИЯ ЧЕЛОВЕЧЕСТВА
У млекопитающих — маммалийный тип мозга. Для него характерно сильное развитие переднего мозга за счет коры, которая развивается на основе небольшого островка коры пресмыкающихся и становится интегрирующим центром мозга. В ней располагаются высшие центры зрительного, слухового, осязательного, двигательного анализаторов, а также центры высшей нервной деятельности. Кора имеет очень сложное строение и называется новой корой — neocortex. В ней располагаются не только тела нейронов, но и ассоциативные волокна, соединяющие разные ее участки. Характерным является также наличие комиссуры между обоими полушариями, в которой располагаются волокна, связывающие их воедино. Промежуточный мозг, как и у других классов, включает гипоталамус, гипофиз и эпифиз. В среднем мозге располагается четверохолмие в виде четырех бугров. Два передних связаны со зрительным анализатором, два задних—со слуховым. Очень хорошо развит мозжечок (рис. 14.39).
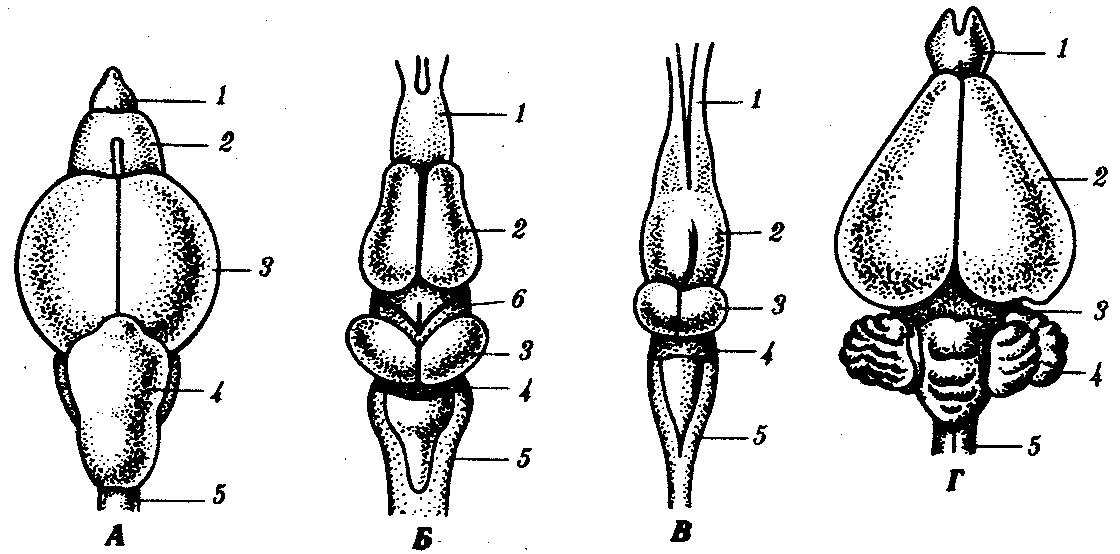
Рис. 14.39. Эволюция головного мозга позвоночных. А—рыба; Б—земноводное; В—пресмыкающееся; Г—млекопитающее:
1—обонятельные доли, 2—передний мозг, 3—средний мозг, 4—мозжечок, 5—продолговатый мозг, 6—промежуточный мозг
По мере усиления функций передних отделов головного мозга в филогенезе спинного мозга наблюдается его продольная дифференцировка с образованием утолщений в области отхождения крупных нервов к конечностям и редукция его заднего конца.
Так, у рыб спинной мозг равномерно тянется вдоль всего тела. Начиная от земноводных происходит его укорочение сзади. У млекопитающих на заднем конце спинного мозга остается рудимент в виде конечной нити — filum terminale. Нервы, идущие к заднему концу тела, проходят по позвоночному каналу самостоятельно, образуя так называемый конский хвост — cauda equina.
Основные этапы эволюции центральной нервной системы отражаются и в онтогенезе человека. На стадии нейруляции закладывается нервная пластинка, превращающаяся в желобок и затем в трубку. Передний конец трубки образует сначала три мозговых пузыря (рис. 14.40): передний (
I), средний (II) и задний (III). Вслед за этим передний пузырь подразделяется на два, дифференцирующихся на передний (7) и промежуточный (2) мозг — telencephalon, diencephalon. Средний мозговой пузырь развивается в средний (3) мозг — mesencephalon, a задний — в задний (4) мозг — metencephalon — и продолговатый (5) мозг — medulla oblongata.
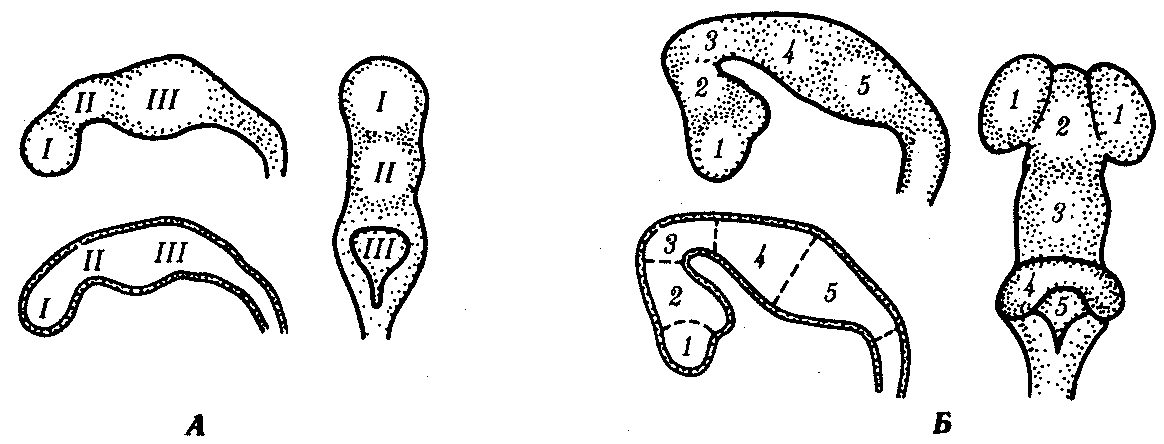
Рис. 14.40. Мозговые пузыри в эмбриогенезе мозга человека. А — стадия трех мозговых пузырей; Б — стадия пяти мозговых пузырей (остальные пояснения см. в тексте)
Задний конец спинного мозга редуцируется, превращаясь в терминальную нить. Позже скорости роста спинного мозга и позвоночника оказываются разными, и к моменту рождения конец спинного мозга оказывается на уровне третьего, а у взрослого человека — уже на уровне первого поясничного позвонка.
Головной мозг рано начинает развиваться по пути, характерному для млекопитающих и человека. Первично почти прямая нервная трубка резко изгибается в области будущего продолговатого и среднего мозга. На этом фоне большие полушария переднего мозга растут с особенно большой скоростью. В результате головной мозг оказывается расположенным над лицевым черепом. Дифференцировка коры приводит к развитию извилин, борозд и формированию высших сенсорных и двигательных центров, в том числе центров письменной и устной речи и др., характерных только для человека (см. § 15.3).
Центральная нервная система столь важна для интеграции индивидуального развития человека, что большинство ее врожденных пороков несовместимы с жизнью. Среди пороков спинного мозга, онтогенетические механизмы которых известны, отметим рахисхиз,нли платиневрию,— отсутствие замыкания нервной трубки (рис. 14.41). Эта аномалия связана с нарушением клеточных перемещений и адгезии в зоне формирования нервной трубки в процессе нейруляции. Аномалия переднего мозга — прозэнцефалия — выражается в нарушении морфо-генеза мозга, при котором полушария оказываются неразделенными, а кора — недоразвита. Этот порок формируется на 4-й неделе эмбриогенеза, в момент закладки переднего мозга. Как и предыдущий, он несовместим с жизнью. Часто встречается у мертворожденных при различных хромосомных и генных синдромах.
Нарушения дифференцировки коры — агирия (отсутствие извилин) и олигогирия с пахигирией (малое количество утолщенных извилин) — сопровождаются упрощением гистологического строения коры. У детей с такими пороками выявляются грубая олигофрения и нарушение многих рефлексов. Большинство детей умирают в течение первого года жизни.


Рис. 14.41. Рахисхиз (платиневрия)
14.6.2. Эндокринная система
Эндокринная система наряду с нервной является ведущим аппаратом интеграции многоклеточного организма, обеспечивая гуморальную регуляцию функций органов. Эта регуляция осуществляется гормонами — биологически активными веществами разной химической природы, выделяемыми железами внутренней секреции. Действие гормонов строго специфично: разные гормоны действуют на разные органы, вызывая определенные изменения их функционирования. Железы внутренней секреции не имеют протоков и выделяют гормоны непосредственно в кровь, что облегчает их транспорт к органам-мишеням. Клетки органов-мишеней на своих мембранах имеют специфические рецепторы, с которыми связываются гормоны, вызывая определенные изменения их метаболизма. Гуморальная регуляция эволюционно возникла значительно раньше нервной потому, что она более проста и не требует развития таких сложных структур, как нервная система.
Обсуждая вопрос о филогенезе эндокринной системы хордовых, необходимо рассмотреть происхождение и эволюцию как гормонов, так и самих желез внутренней секреции.
14.6.2.1. Гормоны
Химическая структура гормонов многообразна. Это могут быть белки (инсулин, пептидные нейрогормоны), стероиды (половые гормоны), продукты метаболизма отдельных аминокислот (тироксин, адреналин). Они могут иметь и другое строение. Это свидетельствует о том, что гормоны разнообразны и по происхождению. Главное, что их объединяет, — способность специфически изменять клеточный метаболизм при контакте с цитоплазматической мембраной.
Предполагают, что уже древние одноклеточные организмы использовали биологически активные вещества для межклеточных коммуникаций. Некоторые вещества такого рода, обладающие регуляторными функциями, могли действовать как на отдельные клетки простейших, так позже и на клетки многоклеточных организмов. Впоследствии они и стали выполнять функции гормонов. Интересно, что в прогрессивной эволюции гуморальной регуляции структура самих гормонов может и не меняться. Доказательством этого является обнаружение таких известных гормонов, как адреналин, норадреналин и некоторые другие, в клетках простейших и низших растений, где они выполняют функции регуляторов клеточного деления, движения ресничек и вакуолей.
В эмбриогенезе многоклеточных ряд гормонов выявляется уже в первые часы и дни развития. В процессе дробления они регулируют течение клеточного цикла. Позже — перемещения клеток и образование межклеточных контактов, действуя либо внутри клеток, их продуцирующих, либо на близлежащие клетки. Гормоны приобретают свойства дистантных регуляторов в филогенезе только у трехслойных животных, а в онтогенезе многоклеточных — соответственно на стадии первичного органогенеза.
При неизменности химической структуры функции гормонов могут нередко изменяться. Так, гормон пролактин, выделяющийся у млекопитающих и человека гипофизом и регулирующий секрецию молока млечными железами, обнаружен также у рыб, земноводных и птиц. У первых он регулирует выделение кожными железами слизи, которой питаются мальки, у вторых — образование оболочек икринок в яйцеводах, у третьих — некоторые элементы брачного поведения, а также выделение у кормящих родителей «зобного молочка». Как видно из этого примера, эволюция действия гормонов может быть канализированной выполнением одной и той же функции в самом широком смысле. Действительно, все перечисленные функции пролактина у животных, находящихся на разных уровнях организации, имеют отношение к обеспечению успешности размножения.
Функции других гормонов, например адреналина, могут в филогенетическом ряду тех же позвоночных практически не меняться, обеспечивая у всех регуляцию в первую очередь энергетического обмена.
При этом часто гормоны продолжают выполнять те функции, которые первично проявлялись еще у одноклеточных и низших многоклеточных животных. Так, адреналин в организме млекопитающих и человека замедляет прохождение клеток по клеточному циклу и вступление их в митоз. Из главной эта функция у высокоорганизованных животных превратилась во второстепенную. Таким образом, на филогенез гормонов, так же как и на эволюцию органов, распространяются основные закономерности макроэволюции (см. гл. 13).
14.6.2.2. Железы внутренней секреции
Железы внутренней секреции, как и гормоны, выделяемые ими, имеют разное происхождение, что важно для изучения их эволюции.
Некоторые эндокринные железы связаны по происхождению с эпителиальной выстилкой глотки. К ним относятся щитовидная и паращитовидная железы. Эпифиз развивается как вырост мозга; гипофиз, надпочечники и поджелудочная железа
имеют сложное происхождение.
Среди хордовых только у бесчерепных эндокринная система существует в виде отдельных клеток и клеточных комплексов, которые находятся в разных отделах тела, объединенных друг с другом за счет гуморального взаимодействия. У позвоночных в основании промежуточного мозга развивается гипоталамус — нейросекреторное образование, осуществляющее связь между двумя системами интеграции организма в единое целое: нервной и эндокринной. Вместе с гипофизом гипоталамус образует единую гипоталамо-гипофизарную систему.
Эволюционным предшественником гипоталамуса является так называемый инфундибулярный вырост ланцетника, состоящий из нейросекреторных клеток и находящийся на вентральной стороне переднего конца нервной трубки.
Начиная с рыб гипоталамус дифференцируется на многочисленные ядра, клетки которых с помощью отростков контактируют как с нейронами мозга, так и с клетками гипофиза. Нейросекреторные клетки гипоталамуса выделяют две основные группы гормонов: пептидные и моноаминовые.
Первые — гормоны, влияющие на функции внутренних органов — вазопрессин, регулирующий артериальное давление, окситоцин, действующий на мускулатуру матки, и др. Вторая группа гормонов (дофамин, норадреналин, серотонин) регулирует деятельность передней доли гипофиза. Под их действием стимулируется или подавляется секреция гормонов соответствующими гипофизарными клетками.
Подробнее остановимся на эволюции гипофиза. Эта железа состоит из трех долей: передней (аденогипофиза), средней (промежуточной) и задней (нейрогипофиза). Гипофиз соединен с гипоталамусом его выростом — воронкой, через которую проходят отростки нейронов гипоталамуса и кровеносные сосуды, обеспечивающие их гуморальное взаимодействие.
Доли гипофиза имеют разное происхождение. Передняя доля развивается из выпячивания эктодермального эпителия крыши ротовой полости, так называемого кармана Ратке, который растет в сторону промежуточного мозга. Задняя доля развивается из задней части воронки. Клетки, входящие в ее состав, по происхождению являются глиальными. Промежуточная доля — производная от передней (рис. 14.42).
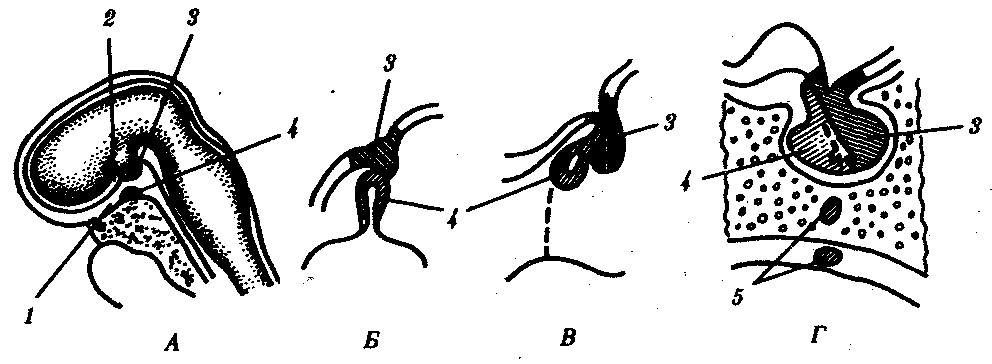
Рис. 14.42. Развитие гипофиза у человека. А—сагиттальный срез 4-недельного зародыша; Б—объединение зачатков гипофиза из стенки глотки и основания мозга; В—формирование трех долей гипофиза на 8-й неделе развития;