ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 03.02.2024
Просмотров: 998
Скачиваний: 0

СОДЕРЖАНИЕ
ПОПУЛЯЦИОННО-ВИДОВОЙ УРОВЕНЬ ОРГАНИЗАЦИИ ЖИЗНИ
ВИДООБРАЗОВАНИЕ В ПРИРОДЕ. ЭЛЕМЕНТАРНЫЕ ЭВОЛЮЦИОННЫЕ ФАКТОРЫ
11.5. ГЕНЕТИКО-АВТОМАТИЧЕСКИЕ ПРОЦЕССЫ (ДРЕЙФ ГЕНОВ)
11.7. НАСЛЕДСТВЕННЫЙ ПОЛИМОРФИЗМ ПРИРОДНЫХ ПОПУЛЯЦИЙ.
11.9. ПРОИСХОЖДЕНИЕ БИОЛОГИЧЕСКОЙ ЦЕЛЕСООБРАЗНОСТИ
ДЕЙСТВИЕ ЭЛЕМЕНТАРНЫХ ЭВОЛЮЦИОННЫХ ФАКТОРОВ
12.1. ПОПУЛЯЦИЯ ЛЮДЕЙ. ДЕМ, ИЗОЛЯТ
12.2. ВЛИЯНИЕ ЭЛЕМЕНТАРНЫХ ЭВОЛЮЦИОННЫХ ФАКТОРОВ НА ГЕНОФОНДЫ ЧЕЛОВЕЧЕСКИХ ПОПУЛЯЦИЙ
12.3. ГЕНЕТИЧЕСКОЕ РАЗНООБРАЗИЕ
13.1. ЭВОЛЮЦИЯ ГРУПП ОРГАНИЗМОВ
13.2. СООТНОШЕНИЕ ОНТО- И ФИЛОГЕНЕЗА
13.4. Организм как целое в историческом
Соотносительные преобразования органов
13.5. СОВРЕМЕННАЯ СИСТЕМА ОРГАНИЧЕСКОГО МИРА
ФИЛОГЕНЕЗ СИСТЕМ ОРГАНОВ ХОРДОВЫХ
14.2. ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
И ДАЛЬНЕЙШАЯ ЭВОЛЮЦИЯ ЧЕЛОВЕКА
15.3. ХАРАКТЕРИСТИКА ОСНОВНЫХ ЭТАПОВ АНТРОПОГЕНЕЗА
15.4. ВНУТРИВИДОВАЯ ДИФФЕРЕНЦИАЦИЯ ЧЕЛОВЕЧЕСТВА
БИОГЕОЦЕНОТИЧЕСКИЙ УРОВЕНЬ ОРГАНИЗАЦИИ ЖИЗНИ
16.1. БИОГЕОЦЕНОЗ - ЭЛЕМЕНТАРНАЯ ЕДИНИЦА БИОГЕОЦЕНОТИЧЕСКОГО УРОВНЯ ОРГАНИЗАЦИИ ЖИЗНИ
17.2. ЧЕЛОВЕК КАК ОБЪЕКТ ДЕЙСТВИЯ ЭКОЛОГИЧЕСКИХ ФАКТОРОВ.
АДАПТАЦИЯ ЧЕЛОВЕКА К СРЕДЕ ОБИТАНИЯ
17.4. РОЛЬ АНТРОПОГЕННЫХ ФАКТОРОВ
В ЭВОЛЮЦИИ ВИДОВ И БИОГЕОЦЕНОЗОВ
МЕДИЦИНСКАЯ ПАРАЗИТОЛОГИЯ. ОБЩИЕ ВОПРОСЫ
18.2. ФОРМЫ МЕЖВИДОВЫХ БИОТИЧЕСКИХ СВЯЗЕЙ В БИОЦЕНОЗАХ
18.3. КЛАССИФИКАЦИЯ ПАРАЗИТИЗМА
18.4. РАСПРОСТРАНЕННОСТЬ ПАРАЗИТИЗМА
18.5. ПРОИСХОЖДЕНИЕ ПАРАЗИТИЗМА
18.6. АДАПТАЦИИ К ПАРАЗИТИЧЕСКОМУ ОБРАЗУ ЖИЗНИ. ОСНОВНЫЕ ТЕНДЕНЦИИ
18.8. ФАКТОРЫ ВОСПРИИМЧИВОСТИ ХОЗЯИНА К ПАРАЗИТУ
18.9. ДЕЙСТВИЕ ХОЗЯИНА НА ПАРАЗИТА
18.10. СОПРОТИВЛЕНИЕ ПАРАЗИТОВ РЕАКЦИЯМ ИММУНИТЕТА ХОЗЯИНА
18.11. ВЗАИМООТНОШЕНИЯ В СИСТЕМЕ ПАРАЗИТ - ХОЗЯИН НА УРОВНЕ ПОПУЛЯЦИЙ
18.12. СПЕЦИФИЧНОСТЬ ПАРАЗИТОВ ПО ОТНОШЕНИЮ К ХОЗЯИНУ
18.13. ПРИРОДНО-ОЧАГОВЫЕ ЗАБОЛЕВАНИЯ
19.2. Простейшие, обитающие в полостных органах, сообщающихся с внешней средой
19.3. Простейшие, обитающие в тканях
19.4. ПРОСТЕЙШИЕ — ФАКУЛЬТАТИВНЫЕ ПАРАЗИТЫ ЧЕЛОВЕКА
20.1. ТИП ПЛОСКИЕ ЧЕРВИ PLATHELMINTHES
20.2. ТИП КРУГЛЫЕ ЧЕРВИ NEMATHELMINTHES
21.1. КЛАСС ПАУКООБРАЗНЫЕ ARACHNOIDEA
ЭВОЛЮЦИЯ ПАРАЗИТОВ И ПАРАЗИТИЗМА ПОД ДЕЙСТВИЕМ АНТРОПОГЕННЫХ ФАКТОРОВ
23.1. ПРОИСХОЖДЕНИЕ ЯДОВИТОСТИ
23.2. ЧЕЛОВЕК И ЯДОВИТЫЕ ЖИВОТНЫЕ
24.1. СОВРЕМЕННЫЕ КОНЦЕПЦИИ БИОСФЕРЫ
24.2. СТРУКТУРА И ФУНКЦИИ БИОСФЕРЫ
25.2. ПУТИ ВОЗДЕЙСТВИЯ ЧЕЛОВЕЧЕСТВА
Волосы многих млекопитающих дифференцированы на типичные, служащие для терморегуляции, и большие, или вибриссы, основания которых связаны с чувствительными нервными окончаниями. У большинства млекопитающих вибриссы расположены в области рта и носа, у приматов они редуцированы в связи с усилением осязательной функции передних конечностей, у многих яйцекладущих и сумчатых — разбросаны по всему телу. Этот факт может свидетельствовать о том, что волосяной покров предков млекопитающих первично выполнял осязательные функции, а затем, по мере увеличения количества волос, стал принимать участие в терморегуляции. В онтогенезе человека закладывается большее количество волосяных зачатков, но к концу эмбриогенеза наступает редукция большинства из них.
 Рис. 14.2. Расположение волос у млекопитающих. А—на хвосте грызунов; Б—на других участках тела: 1—роговые чешуйки, 2—группы волос, расположенные в шахматном порядке | 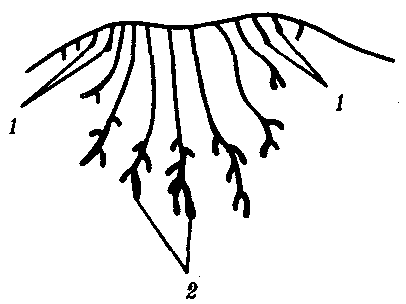 Рис. 14.3. Строение развивающегося соска млекопитающего Виден постепенный переход от потовых (1) к млечным (2) железам |
Потовые железы млекопитающих гомологичны кожным железам амфибий. Их секрет может быть слизистым, содержать белки и жир. Некоторые потовые железы дифференцировались у ранних млекопитающих в млечные железы. У яйцекладущих (утконос, ехидна) млечные железы сходны с потовыми по строению и развитию. По краям развивающегося соска молочной железы можно обнаружить последовательные переходы от типичных потовых к млечным железам (рис. 14.3). Количество млечных желез и сосков коррелирует с плодовитостью (от 25 до одной пары), но в эмбриогенезе всех млекопитающих на брюшной поверхности закладываются «млечные линии», тянущиеся от подмышечной впадины до паха. Впоследствии на этих линиях дифференцируются соски, большая часть которых затем подвергается редукции и исчезает. Так, в эмбриогенезе человека закладывается вначале пять пар сосков, а впоследствии остается лишь одна (рис. 14.4).

Рис. 14.4. Эмбриогенез передней брюшной стенки человека. А — зародыш в возрасте 5 недель (видны млечные линии); Б — дифференцировка пяти пар сосков; В — зародыш в возрасте 7 недель
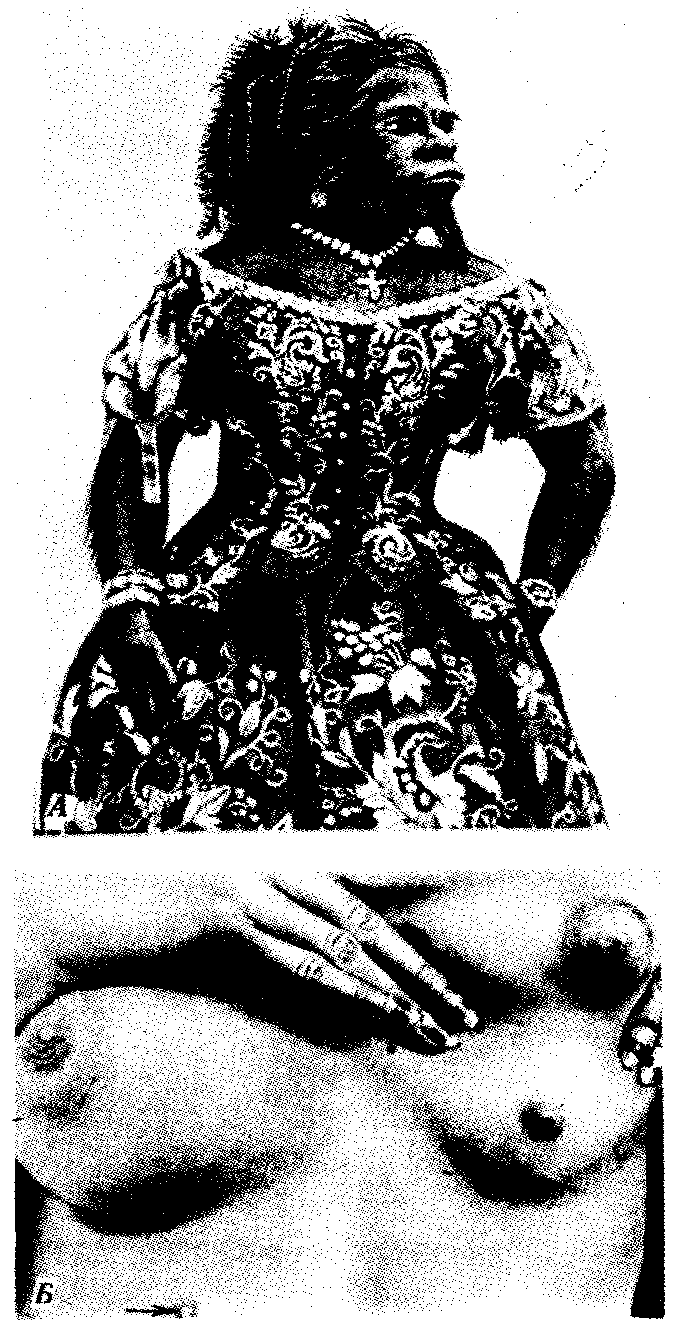
Рис. 14.5. Атавистические аномалии развития кожи.
А — гипертрихоз; Б — полимастия
Сальные железы образуются в коже только у млекопитающих. Их секрет, смазывая волосы и поверхность кожи, придает им несмачиваемость и эластичность.
Онтогенез покровов и придатков кожи млекопитающих и человека отражает их эволюцию по типу архаллаксиса. Действительно, ни зачатки роговых чешуи, характерных для пресмыкающихся, ни более ранние формы придатков кожи в их эмбриогенезе не рекапитулируют. При этом на стадии вторичного органогенеза развиваются сразу зачатки волосяных фолликулов. Нарушения раннего онтогенеза кожных покровов человека могут вызвать возникновение некоторых малосущественных атавистических пороков развития: гипертрихоз (повышенное оволосение), полителию (увеличенное количество сосков), полимастию (увеличенное количество млечных желез) (рис. 14.5). Все они связаны с нарушением редукции избыточного количества этих структур и отражают эволюционную связь человека с наиболее близкими предковыми формами — млекопитающими. Именно поэтому у человека и других млекопитающих невозможно рождение потомства с атавистическими признаками кожных покровов, характерными для более отдаленных предков. Одним из самых известных признаков недоношенности новорожденных является повышенное оволосение кожи. Вскоре после рождения избыточные волосы обычно выпадают, а их фолликулы редуцируются.

14.2. ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
Филогенез двигательной функции лежит в основе прогрессивной эволюции животных. Поэтому уровень их организации в первую очередь зависит от характера двигательной активности, которая определяется особенностями организации опорно-двигательного аппарата, претерпевшего в типе Хордовые большие эволюционные преобразования в связи со сменой сред обитания и изменения форм локомоции. Действительно, водная среда у животных, не имеющих наружного скелета, предполагает однообразные движения за счет изгибов всего тела, в то время как жизнь на суше более способствует их перемещению с помощью конечностей.
Рассмотрим в отдельности эволюцию скелета и мышечной системы.
14.2.1. Скелет
У хордовых скелет внутренний. По строению и функциям подразделяется на осевой, скелет конечностей и головы.
14.2.1.1. Осевой скелет
В подтипе Бесчерепные имеется только осевой скелет в виде хорды. Она построена из сильно вакуолизированных клеток, плотно прилегающих друг к другу и покрытых снаружи общими эластической и волокнистой оболочками. Упругость хорде придают тургорное давление ее клеток и прочность оболочек. Хорда закладывается в онтогенезе всех хордовых и выполняет у более высокоорганизованных животных не столько опорную, сколько морфогенетическую функцию, являясь органом, осуществляющим эмбриональную индукцию.
На протяжении всей жизни у позвоночных хорда сохраняется только у круглоротых и некоторых низших рыб. У всех остальных животных она редуцируется. У человека в постэмбриональном периоде сохраняются рудименты хорды в виде nucleus pulposus межпозвоночных дисков. Сохранение избыточного количества хордального материала при нарушении его редукции чревато возможностью развития у человека опухолей — хордом, возникающих на его основе.
У всех позвоночных хорда постепенно вытесняется позвонками, развивающимися из склеротомов сомитов, и функционально заменяется позвоночным столбом. Это один из выраженных примеров гомотопной субституции органов (см. § 13.4). Формирование позвонков в филогенезе начинается с развития их дуг, охватывающих нервную трубку и становящихся местами прикрепления мышц. Начиная с хрящевых рыб обнаруживается охрящевение оболочки хорды и разрастание оснований позвонковых дуг, в результате чего формируются тела позвонков. Срастание верхних позвонковых дуг над нервной трубкой образует остистые отростки и позвоночный канал, в который заключена нервная трубка (рис. 14.6).
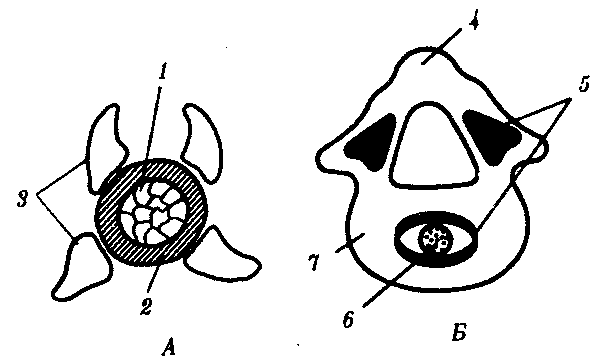
Рис. 14.6. Развитие позвонка. А—ранний этап; Б—последующая стадия:
1—хорда, 2—оболочка хорды, 3—верхние и нижние позвонковые дуги, 4—остистый отросток, 5—зоны окостенения, 6—рудимент хорды, 7—хрящевое тело позвонка
Замещение хорды позвоночным столбом — более мощным органом опоры, имеющим сегментарное строение,— позволяет увеличить общие размеры тела и активизирует двигательную функцию. Дальнейшие прогрессивные изменения позвоночного столба связаны с тканевой субституцией — заменой хрящевой ткани на костную, что обнаруживается у костных рыб, а также с дифференцировкой его на отделы.
У рыб только два отдела позвоночника: туловищный и хвостовой. Это связано с перемещением их в воде за счет изгибов тела.
Земноводные приобретают также шейный и крестцовый отделы, представленные каждый одним позвонком. Первый обеспечивает большую подвижность головы, а второй — опору задним конечностям.
У пресмыкающихся удлиняется шейный отдел позвоночника, первые два позвонка которого подвижно соединены с черепом и обеспечивают большую подвижность головы. Появляется поясничный отдел, еще слабо отграниченный от грудного, а крестец состоит уже из двух позвонков.
Млекопитающие характеризуются стабильным количеством позвонков в шейном отделе, равным 7. В связи с большим значением в движении задних конечностей крестец образован 5—10 позвонками. Поясничный и грудной отделы четко отграничены друг от друга.
У рыб все туловищные позвонки несут ребра, не срастающиеся друг с другом и с грудиной. Они придают телу устойчивую форму и обеспечивают опору мышцам, изгибающим тело в горизонтальной плоскости. Эта функция ребер сохраняется у всех позвоночных, совершающих змеевидные движения,— у хвостатых земноводных и пресмыкающихся , поэтому у них ребра также располагаются на всех позвонках, кроме хвостовых.
У пресмыкающихся часть ребер грудного отдела срастается с грудиной, формируя грудную клетку, а у млекопитающих в состав грудной клетки входит 12—13 пар ребер.
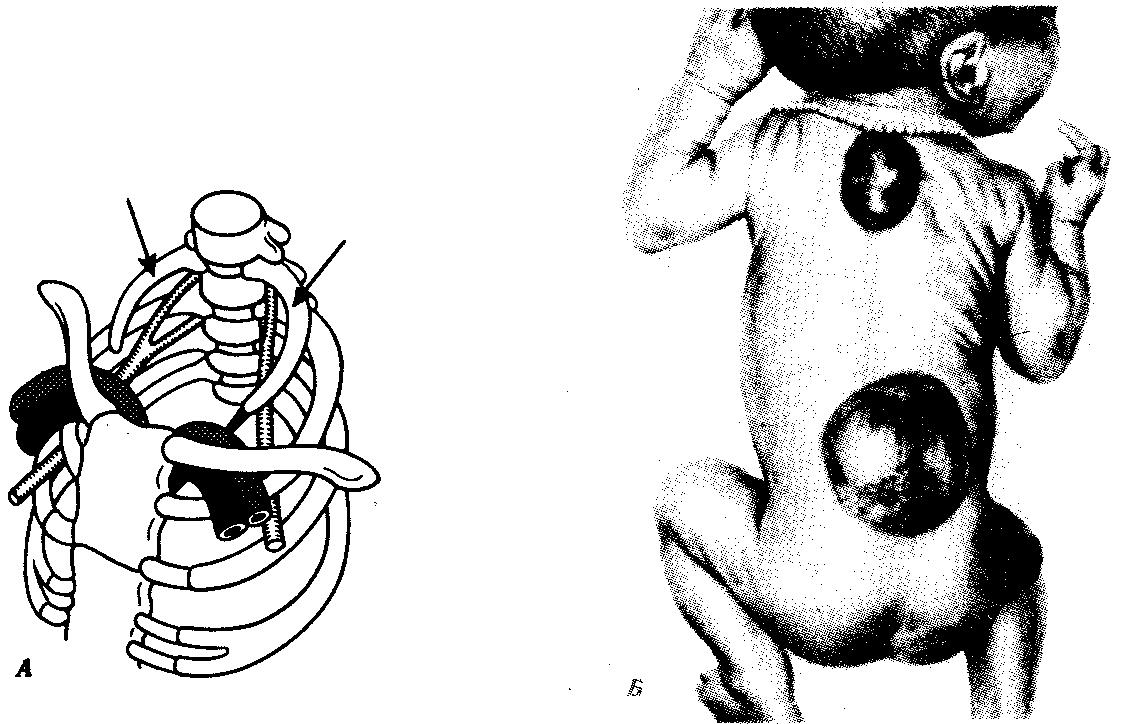
Рис. 14.7. Аномалии развития осевого скелета. А — рудиментарные шейные ребра (показаны стрелками); Б — несращение остистых отростков позвонков в грудной и поясничной областях. Спинномозговые грыжи
Онтогенез осевого скелета человека рекапитулирует основные филогенетические стадии его становления: в периоде нейруляции закладывается хорда, заменяющаяся впоследствии хрящевым, а затем и костным позвоночником. На шейных, грудных и поясничных позвонках развивается по паре ребер, после чего шейные и поясничные ребра редуцируются, а грудные срастаются спереди друг с другом и с грудиной, формируя грудную клетку.
Нарушение онтогенеза осевого скелета у человека может выразиться в таких атавистических пороках развития, как несрастание остистых отростков позвонков, в результате чего формируется spinabifida — дефект позвоночного канала. При этом часто через дефект выпячиваются мозговые оболочки и образуется спинномозговая грыжа (рис. 14.7).
В возрасте 1,5—3 мес. зародыш человека обладает хвостовым отделом позвоночника, состоящим из 8—11 позвонков. Нарушение их редукции в последующем объясняет возможность возникновения такой известной аномалии осевого скелета, как персистирование хвоста.
Нарушение редукции шейных и поясничных ребер лежит в основе их сохранения в постнатальном онтогенезе.
14.2.1.2. Скелет головы
Продолжением осевого скелета спереди является осевой, или мозговой, череп, служащий для защиты головного мозга и органов чувств. Рядом с ним развивается висцеральный, или лицевой череп, образующий опору передней части пищеварительной трубки. Обе части черепа развиваются по-разному и из разных зачатков. На ранних этапах эволюции и онтогенеза они не связаны между собой, но позже эта связь возникает.
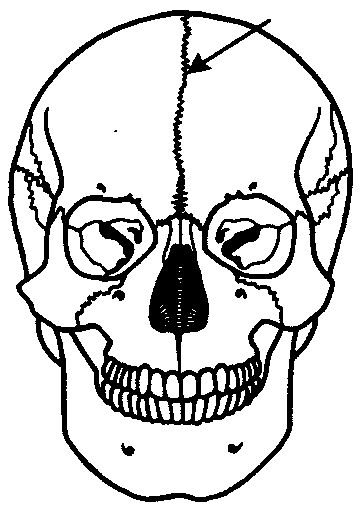
Рис. 14.8. Череп человека с методическим швом (указан стрелкой)
В задней части осевого черепа в процессе развития обнаруживаются следы сегментации, поэтому считают, что он представляет собой результат слияния друг с другом закладок передних позвонков. В состав мозгового черепа включаются также закладки хрящевых капсул мезенхимального происхождения, окружающие органы слуха, обоняния и зрения. Кроме того, часть мозгового черепа (лежащая кпереди от турецкого седла), не имеющая сегментации, развивается, по-видимому, как новообразование в связи с увеличением размеров переднего мозга.
Филогенетически мозговой череп прошел три стадии развития: перепончатую, хрящевую и костную.
У круглоротых он практически весь перепончатый и не имеет передней, несегментированной, части.