Файл: Тема 1 клетка Вопрос Химический состав, организация плазмолеммы. Функции плазмолеммы.docx
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 17.03.2024
Просмотров: 317
Скачиваний: 0
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.

, образованной высокими кубическими клетками. Большинство из них относится к светлым, или главным, среди которых рассеяны одиночные вставочные клетки (тёмные). Со стороны просвета собирательной трубочки главные клетки имеют гладкую поверхность и одну (иногда две) ресничку. Апикальная мембрана вставочных клеток образует множество мелких уплощённых выростов. В цитоплазме этих клеток содержится много митохондрий. По мере увеличения калибра собирательных трубочек вставочные клетки постепенно исчезают. Количество тёмных клеток в собирательных трубочках значительно увеличивается при респираторном ацидозе.
В собирательных трубочках происходит завершающий этап реабсорбции воды и концентрирование мочи. Вода выходит из трубочек через водные каналы, образованные аквапоринами 2 и 3.
Аквапорин 2 содержится в везикулах апикальной части главных клеток. Аквапорин 2‑водные каналы регулируются вазопрессином (АДГ). При повышенном содержании натрия в крови (гипернатриемии) или пониженном объёме крови (гиповолемии) усиливается поступление АДГ в кровь. АДГ связывается с рецепторами вазопрессина V2 в базолатеральной мембране главных клеток. Это стимулирует выход аквапорина 2 из везикул и образование дополнительных водных каналов в апикальной мембране клетки. После отделения АДГ от своего рецептора аквапорин 2 возвращается в цитоплазму путём эндоцитоза. Таким образом, вазопрессин способствует задержке в организме воды, уменьшению диуреза и увеличению АД. С другой стороны, вазопрессин прямо влияет на ГМК сосудистой стенки (через рецепторы V1a), вызывая сосудосуживающий эффект.
Аквапорин 3 образует водный канал в базолатеральной плазмолемме клеток собирательных трубочек мозгового вещества.
Вопрос 11. Юкстагломерулярный аппарат нефрона. Ренин и регуляция артериального давления.
Юкстагломерулярный комплекс – околоклубочковый комплекс, регулирующий функцию нефрона. В юкстагломерулярном комплексе различают плотное пятно, юкстагломерулярные и юкставаскулярные клетки.
Плотное пятно
Плотное пятно образовано клетками дистального извитого канальца в области его перегиба между приносящей и выносящей артериолами клубочка. Клетки плотного пятна имеют сравнительно большую высоту, ядра смещены к основанию клеток. Эти клетки тесно контактируют с юкстагломерулярными и юкставаскулярными клетками, поскольку в этом участке канальца отсутствует базальная мембрана.
Юкстагломерулярные клетки — видоизменённые ГМК средней оболочки приносящей артериолы. Они имеют полигональную форму и округлое, а не вытянутое, как у ГМК, ядро. У юкстагломерулярных клеток хорошо развиты гранулярная эндоплазматическая сеть и комплекс Гольджи. В цитоплазме отсутствуют характерные для ГМК филаменты, но имеется большое количество как отдельных, так и образующих агрегаты секреторных гранул, содержащих ренин.
Юкставаскулярные клетки образуют скопление (полюсную подушку) между плотным пятном и клубочком в углублении между приносящей и выносящей артериолами. Для клеток полюсной подушки характерны неправильная форма, бледное ядро, филаменты в цитоплазме, а также длинные отростки. К настоящему времени функция клеток остаётся неустановленной. Считают, что клетки полюсной подушки содержат ангиотензиназу А. Экстрагломерулярные мезангиальные клетки, по некоторым данным, могут участвовать в синтезе ренина при истощении функции юкстагломерулярных клеток.
Ренин накапливается в секреторных гранулах и при наличии соответствующего сигнала секретируется в просвет приносящей артериолы. В соответствии с барорецепторной теорией, ренин-синтезирующие клетки функционируют как барорецепторы. Повышение давления в просвете приносящей артериолы снижает секрецию ренина. При снижении давления уменьшается растяжение стенки артериолы, что вызывает усиление секреции ренина юкстагломерулярными клетками. Выделение норадреналина из многочисленных окончаний аксонов симпатических нейронов в области околоклубочкового комплекса увеличивает секрецию ренина. Клетки плотного пятна регистрируют содержание ионов Na+ в просвете дистального канальца. При избыточном содержании ионов Na+ (в этом случае возрастает осмотическое давление в канальце) секреция ренина юкстагломерулярными клетками ингибируется.
Вопрос 12. Строение оболочек мочевого пузыря, мочеточников. Особенности строения переходного эпителия.
Переходный эпителий. Клетки поверхностного слоя переходного эпителия имеют округлую или куполообразную форму. Плазмолемма апикальной части этих клеток содержит специальные пластинки полигональной формы, благодаря чему на поверхности клеток формируется разветвлённая сеть микроскладок. Складки расправляются по границам между пластинками при растяжении стенки органа, так что клетки уплощаются и вытягиваются, а эпителий в целом становится тоньше. Между клетками эпителия формируются плотные контакты, что предотвращает проникновение содержимого в подлежащую ткань.

Мочеточник.
Стенка мочеточника состоит из слизистой, подслизистой, мышечной и наружной оболочек. Слизистая оболочка мочеточника собрана в продольные складки. Переходный эпителий состоит из 6–8 слоёв клеток. В собственном слое слизистой оболочки, представленном соединительной тканью, присутствуют в основном коллагеновые и отдельные эластические волокна, изредка встречаются лимфатические фолликулы. Мышечный слой слизистой оболочки отсутствует. Вблизи мышечной оболочки собственный слой слизистой оболочки становится более рыхлым; эту часть иногда выделяют в отдельную оболочку — подслизистую. В подслизистой оболочке нижней части мочеточника имеются мелкие альвеолярно-трубчатые железы. Мышечная оболочка в верхних двух третях мочеточника состоит из двух слоёв ГМК: внутреннего (продольного) и наружного (циркулярного); в нижней трети снаружи добавляется третий слой (продольный). Адвентициальная оболочка представлена волокнистой соединительной тканью, содержащей много эластических волокон. По периферии оболочка сливается с прилежащей рыхлой соединительной тканью.
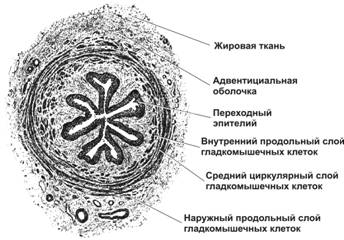
МОЧЕВОЙ ПУЗЫРЬ
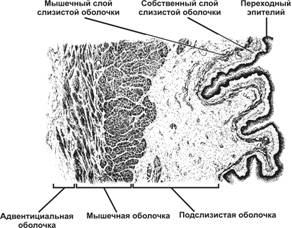
Стенка мочевого пузыря состоит из трёх оболочек: слизистой, мышечной и наружной соединительнотканной. Слизистая оболочка образована наиболее высоким переходным эпителием и собственным слоем. Мышечный слой слизистой оболочки плохо выражен (или вообще отсутствует) и представлен немногочисленными ГМК. Собственный слой слизистой оболочки содержит большое количество коллагеновых и единичные эластические волокна. Наружная его часть состоит из более рыхлой ткани с высоким содержанием эластических волокон; иногда её также выделяют в отдельную оболочку — подслизистую. Последняя, а также сокращение подлежащих слоёв ГМК обусловливают складчатость слизистой оболочки. Мышечная оболочка состоит из трёх слоёв; в среднем из них большинство ГМК имеет циркулярный ход, в наружном и внутреннем — продольный. Адвентициальная оболочка также характеризуется обилием эластических волокон. На верхнезадней поверхности органа она замещается серозной оболочкой.
Тема 20: «Мужская половая система»
Вопрос 1. Развитие мужской половой системы. Первичные гоноциты, начальная локализация, пути миграции в зачаток гонады. Половая дифференцировка.
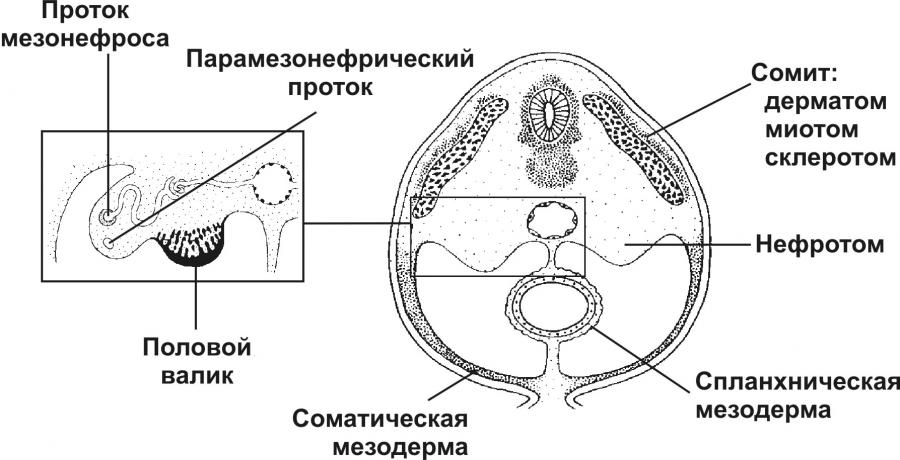
Источниками половых желёз являются урогенитальные валики и первичные половые клетки. Урогенитальные (гонадные) валики — индифферентные гонады — зачатки будущих половых желёз (яичек и яичников). На 4-й неделе эмбриогенеза в грудопоясничном отделе нефротома (на медиальной стороне мезонефроса) формируются гонадные валики, не идентифицируемые ни как мужские, ни как женские половые железы. Индифферентные гонады состоят из заселяемого первичными половыми клетками коркового и мозгового вещества.
Первичные половые клетки возникают, на 2-й неделе эмбрионального развития из клеток головного отдела эпибласта. В ходе гаструляции первичные половые клетки через первичную полоску в энтодерму желточного мешка и далее в гонадные валики. В плодном периоде первичные половые клетки дифференцируются в овогонии в развивающихся яичниках или в сперматогонии в яичках. На пути от овогоний или сперматогоний до зрелых гамет различают несколько стадий: размножения, роста, созревания и формирования.
ПОЛОВАЯ ДИФФЕРЕНЦИРОВКА
Хромосомная детерминация пола происходит при оплодотворении, Y‑хромосома — потенциальная детерминанта генетически мужского пола
Фактор, детерминирующий развитие мужских гонад (TDF) — один из индукторов развития мужской половой железы. Регуляторный фактор TDF, кодируемый геном Y-хромосомы SRY (Sex-determining Region Y), ответственен за дифференцировку яичек из изначально бипотентных зачатков гонад.
Индифферентные гонады
Критическая стадия развития индифферентных гонад — 8-я неделя внутриутробного развития. До 45–50 дня зачатки гонад не имеют половой дифференцировки. Под влиянием фактора транскрипции TDF гонадные валики развиваются как яички; при отсутствии эффектов этого фактора развиваются яичники. Дифференцировку других структур определяют мужские половые гормоны и мюллеров ингибирующий фактор (MIF ¾ Mullerian Inhibiting Factor), продуцируемые в яичках плода.
Дифференцировка по мужскому типу. При кариотипе 46XY из клеток мезенхимы гонадного валика дифференцируются интерстициальные эндокринные клетки яичек плода. Под контролем гонадотропинов (хорионического и гипофизарного) интерстициальные эндокринные клетки яичек плода секретируют тестостерон. Экспрессия гена SRY в поддерживающих эпителиальных клетках инициирует транскрипцию гена, кодирующего MIF.
Дифференцировка по женскому типу при кариотипе 46ХХ происходит при отсутствии определяющего развитие яичек фактора Y-хромосомы, андрогенов и MIF. Гонадные валики развиваются как яичники автономно, под влиянием гормонов плаценты.
Яичник и яичко
Дифференцировка яичника и яичка из индифферентных гонад приведена на рисунке. В женском организме в индифферентных гонадах развивается преимущественно корковое и атрофируется мозговое вещество. В мужском организме преимущественное развитие получает мозговое вещество индифферентной гонады. На 8-й неделе эмбриогенеза яички располагаются на уровне верхних поясничных позвонков. От нижнего полюса яичка вниз тянется поддерживающая связка, выполняющая функцию проводника яичка из брюшной полости через паховый канал в мошонку. Опущение яичек в мошонку завершается приблизительно за 1 месяц до рождения и не происходит при крипторхизме.
Вопрос 2. Яичко. Развитие и строение. Канальцы яичка.
В мужском организме преимущественное развитие получает мозговое вещество индифферентной гонады. На 8-й неделе эмбриогенеза яички располагаются на уровне верхних поясничных позвонков. От нижнего полюса яичка вниз тянется поддерживающая связка, выполняющая функцию проводника яичка из брюшной полости через паховый канал в мошонку. Опущение яичек в мошонку завершается приблизительно за 1 месяц до рождения и не происходит при крипторхизме.
Яички (testis), в отличие от яичников, располагаются вне полости тела (в мошонке). Это обстоятельство важно для нормального течения сперматогенеза, происходящего при температуре 34 °C и ниже. Снаружи яичко покрыто соединительнотканной белочной оболочкой (t. albuginea). Внутренний её слой богат кровеносными сосудами — сосудистая оболочка (t. vasculosa). Утолщение белочной оболочки, вдающееся с одной стороны в паренхиму яичка, называется средостением (mediastinum testis). От средостения внутрь яичка отходят перегородки, разделяющие его на дольки конической формы. Каждая долька содержит от одного до четырёх извитых семенных канальцев (tubulus seminifer contortus), выстланных сперматогенным эпителием (epithelium spermatogenicum). В рыхлой соединительной ткани между семенными канальцами расположены интерстициальные эндокринные клетки (endocrinocytus interstitialis). По мере приближения к средостению семенные канальцы продолжаются в прямые (tubuli recti). Их стенка образована кубическим эпителием и базальной мембраной, окружённой тонкой соединительнотканной оболочкой. Прямые канальцы впадают в сеть яичка (rete testis) — систему анастомозирующих тонкостенных трубочек, продолжающихся в выносящие канальцы придатка. Эпителий, выстилающий сеть яичка, представлен кубическими клетками. Некоторые клетки на апикальной поверхности имеют по одной ресничке, способствующей продвижению сперматозоидов. Под базальной мембраной эпителия располагается рыхлая соединительная ткань, окружающая трубочки rete testis снаружи. Генеративная функция (сперматогенез, spermatogenesis) осуществляется в извитых семенных канальцах, а эндокринная (синтез мужских половых гормонов) — в интерстициальных эндокринных клетках.
В собирательных трубочках происходит завершающий этап реабсорбции воды и концентрирование мочи. Вода выходит из трубочек через водные каналы, образованные аквапоринами 2 и 3.
Аквапорин 2 содержится в везикулах апикальной части главных клеток. Аквапорин 2‑водные каналы регулируются вазопрессином (АДГ). При повышенном содержании натрия в крови (гипернатриемии) или пониженном объёме крови (гиповолемии) усиливается поступление АДГ в кровь. АДГ связывается с рецепторами вазопрессина V2 в базолатеральной мембране главных клеток. Это стимулирует выход аквапорина 2 из везикул и образование дополнительных водных каналов в апикальной мембране клетки. После отделения АДГ от своего рецептора аквапорин 2 возвращается в цитоплазму путём эндоцитоза. Таким образом, вазопрессин способствует задержке в организме воды, уменьшению диуреза и увеличению АД. С другой стороны, вазопрессин прямо влияет на ГМК сосудистой стенки (через рецепторы V1a), вызывая сосудосуживающий эффект.
Аквапорин 3 образует водный канал в базолатеральной плазмолемме клеток собирательных трубочек мозгового вещества.
Вопрос 11. Юкстагломерулярный аппарат нефрона. Ренин и регуляция артериального давления.
Юкстагломерулярный комплекс – околоклубочковый комплекс, регулирующий функцию нефрона. В юкстагломерулярном комплексе различают плотное пятно, юкстагломерулярные и юкставаскулярные клетки.
Плотное пятно
Плотное пятно образовано клетками дистального извитого канальца в области его перегиба между приносящей и выносящей артериолами клубочка. Клетки плотного пятна имеют сравнительно большую высоту, ядра смещены к основанию клеток. Эти клетки тесно контактируют с юкстагломерулярными и юкставаскулярными клетками, поскольку в этом участке канальца отсутствует базальная мембрана.
Юкстагломерулярные клетки — видоизменённые ГМК средней оболочки приносящей артериолы. Они имеют полигональную форму и округлое, а не вытянутое, как у ГМК, ядро. У юкстагломерулярных клеток хорошо развиты гранулярная эндоплазматическая сеть и комплекс Гольджи. В цитоплазме отсутствуют характерные для ГМК филаменты, но имеется большое количество как отдельных, так и образующих агрегаты секреторных гранул, содержащих ренин.
Юкставаскулярные клетки образуют скопление (полюсную подушку) между плотным пятном и клубочком в углублении между приносящей и выносящей артериолами. Для клеток полюсной подушки характерны неправильная форма, бледное ядро, филаменты в цитоплазме, а также длинные отростки. К настоящему времени функция клеток остаётся неустановленной. Считают, что клетки полюсной подушки содержат ангиотензиназу А. Экстрагломерулярные мезангиальные клетки, по некоторым данным, могут участвовать в синтезе ренина при истощении функции юкстагломерулярных клеток.
Ренин накапливается в секреторных гранулах и при наличии соответствующего сигнала секретируется в просвет приносящей артериолы. В соответствии с барорецепторной теорией, ренин-синтезирующие клетки функционируют как барорецепторы. Повышение давления в просвете приносящей артериолы снижает секрецию ренина. При снижении давления уменьшается растяжение стенки артериолы, что вызывает усиление секреции ренина юкстагломерулярными клетками. Выделение норадреналина из многочисленных окончаний аксонов симпатических нейронов в области околоклубочкового комплекса увеличивает секрецию ренина. Клетки плотного пятна регистрируют содержание ионов Na+ в просвете дистального канальца. При избыточном содержании ионов Na+ (в этом случае возрастает осмотическое давление в канальце) секреция ренина юкстагломерулярными клетками ингибируется.
Вопрос 12. Строение оболочек мочевого пузыря, мочеточников. Особенности строения переходного эпителия.
Переходный эпителий. Клетки поверхностного слоя переходного эпителия имеют округлую или куполообразную форму. Плазмолемма апикальной части этих клеток содержит специальные пластинки полигональной формы, благодаря чему на поверхности клеток формируется разветвлённая сеть микроскладок. Складки расправляются по границам между пластинками при растяжении стенки органа, так что клетки уплощаются и вытягиваются, а эпителий в целом становится тоньше. Между клетками эпителия формируются плотные контакты, что предотвращает проникновение содержимого в подлежащую ткань.
Мочеточник.
Стенка мочеточника состоит из слизистой, подслизистой, мышечной и наружной оболочек. Слизистая оболочка мочеточника собрана в продольные складки. Переходный эпителий состоит из 6–8 слоёв клеток. В собственном слое слизистой оболочки, представленном соединительной тканью, присутствуют в основном коллагеновые и отдельные эластические волокна, изредка встречаются лимфатические фолликулы. Мышечный слой слизистой оболочки отсутствует. Вблизи мышечной оболочки собственный слой слизистой оболочки становится более рыхлым; эту часть иногда выделяют в отдельную оболочку — подслизистую. В подслизистой оболочке нижней части мочеточника имеются мелкие альвеолярно-трубчатые железы. Мышечная оболочка в верхних двух третях мочеточника состоит из двух слоёв ГМК: внутреннего (продольного) и наружного (циркулярного); в нижней трети снаружи добавляется третий слой (продольный). Адвентициальная оболочка представлена волокнистой соединительной тканью, содержащей много эластических волокон. По периферии оболочка сливается с прилежащей рыхлой соединительной тканью.
МОЧЕВОЙ ПУЗЫРЬ
Стенка мочевого пузыря состоит из трёх оболочек: слизистой, мышечной и наружной соединительнотканной. Слизистая оболочка образована наиболее высоким переходным эпителием и собственным слоем. Мышечный слой слизистой оболочки плохо выражен (или вообще отсутствует) и представлен немногочисленными ГМК. Собственный слой слизистой оболочки содержит большое количество коллагеновых и единичные эластические волокна. Наружная его часть состоит из более рыхлой ткани с высоким содержанием эластических волокон; иногда её также выделяют в отдельную оболочку — подслизистую. Последняя, а также сокращение подлежащих слоёв ГМК обусловливают складчатость слизистой оболочки. Мышечная оболочка состоит из трёх слоёв; в среднем из них большинство ГМК имеет циркулярный ход, в наружном и внутреннем — продольный. Адвентициальная оболочка также характеризуется обилием эластических волокон. На верхнезадней поверхности органа она замещается серозной оболочкой.
Тема 20: «Мужская половая система»
Вопрос 1. Развитие мужской половой системы. Первичные гоноциты, начальная локализация, пути миграции в зачаток гонады. Половая дифференцировка.
Источниками половых желёз являются урогенитальные валики и первичные половые клетки. Урогенитальные (гонадные) валики — индифферентные гонады — зачатки будущих половых желёз (яичек и яичников). На 4-й неделе эмбриогенеза в грудопоясничном отделе нефротома (на медиальной стороне мезонефроса) формируются гонадные валики, не идентифицируемые ни как мужские, ни как женские половые железы. Индифферентные гонады состоят из заселяемого первичными половыми клетками коркового и мозгового вещества.
Первичные половые клетки возникают, на 2-й неделе эмбрионального развития из клеток головного отдела эпибласта. В ходе гаструляции первичные половые клетки через первичную полоску в энтодерму желточного мешка и далее в гонадные валики. В плодном периоде первичные половые клетки дифференцируются в овогонии в развивающихся яичниках или в сперматогонии в яичках. На пути от овогоний или сперматогоний до зрелых гамет различают несколько стадий: размножения, роста, созревания и формирования.
ПОЛОВАЯ ДИФФЕРЕНЦИРОВКА
Хромосомная детерминация пола происходит при оплодотворении, Y‑хромосома — потенциальная детерминанта генетически мужского пола
Фактор, детерминирующий развитие мужских гонад (TDF) — один из индукторов развития мужской половой железы. Регуляторный фактор TDF, кодируемый геном Y-хромосомы SRY (Sex-determining Region Y), ответственен за дифференцировку яичек из изначально бипотентных зачатков гонад.
Индифферентные гонады
Критическая стадия развития индифферентных гонад — 8-я неделя внутриутробного развития. До 45–50 дня зачатки гонад не имеют половой дифференцировки. Под влиянием фактора транскрипции TDF гонадные валики развиваются как яички; при отсутствии эффектов этого фактора развиваются яичники. Дифференцировку других структур определяют мужские половые гормоны и мюллеров ингибирующий фактор (MIF ¾ Mullerian Inhibiting Factor), продуцируемые в яичках плода.
Дифференцировка по мужскому типу. При кариотипе 46XY из клеток мезенхимы гонадного валика дифференцируются интерстициальные эндокринные клетки яичек плода. Под контролем гонадотропинов (хорионического и гипофизарного) интерстициальные эндокринные клетки яичек плода секретируют тестостерон. Экспрессия гена SRY в поддерживающих эпителиальных клетках инициирует транскрипцию гена, кодирующего MIF.
Дифференцировка по женскому типу при кариотипе 46ХХ происходит при отсутствии определяющего развитие яичек фактора Y-хромосомы, андрогенов и MIF. Гонадные валики развиваются как яичники автономно, под влиянием гормонов плаценты.
Яичник и яичко
Дифференцировка яичника и яичка из индифферентных гонад приведена на рисунке. В женском организме в индифферентных гонадах развивается преимущественно корковое и атрофируется мозговое вещество. В мужском организме преимущественное развитие получает мозговое вещество индифферентной гонады. На 8-й неделе эмбриогенеза яички располагаются на уровне верхних поясничных позвонков. От нижнего полюса яичка вниз тянется поддерживающая связка, выполняющая функцию проводника яичка из брюшной полости через паховый канал в мошонку. Опущение яичек в мошонку завершается приблизительно за 1 месяц до рождения и не происходит при крипторхизме.
Вопрос 2. Яичко. Развитие и строение. Канальцы яичка.
В мужском организме преимущественное развитие получает мозговое вещество индифферентной гонады. На 8-й неделе эмбриогенеза яички располагаются на уровне верхних поясничных позвонков. От нижнего полюса яичка вниз тянется поддерживающая связка, выполняющая функцию проводника яичка из брюшной полости через паховый канал в мошонку. Опущение яичек в мошонку завершается приблизительно за 1 месяц до рождения и не происходит при крипторхизме.
Яички (testis), в отличие от яичников, располагаются вне полости тела (в мошонке). Это обстоятельство важно для нормального течения сперматогенеза, происходящего при температуре 34 °C и ниже. Снаружи яичко покрыто соединительнотканной белочной оболочкой (t. albuginea). Внутренний её слой богат кровеносными сосудами — сосудистая оболочка (t. vasculosa). Утолщение белочной оболочки, вдающееся с одной стороны в паренхиму яичка, называется средостением (mediastinum testis). От средостения внутрь яичка отходят перегородки, разделяющие его на дольки конической формы. Каждая долька содержит от одного до четырёх извитых семенных канальцев (tubulus seminifer contortus), выстланных сперматогенным эпителием (epithelium spermatogenicum). В рыхлой соединительной ткани между семенными канальцами расположены интерстициальные эндокринные клетки (endocrinocytus interstitialis). По мере приближения к средостению семенные канальцы продолжаются в прямые (tubuli recti). Их стенка образована кубическим эпителием и базальной мембраной, окружённой тонкой соединительнотканной оболочкой. Прямые канальцы впадают в сеть яичка (rete testis) — систему анастомозирующих тонкостенных трубочек, продолжающихся в выносящие канальцы придатка. Эпителий, выстилающий сеть яичка, представлен кубическими клетками. Некоторые клетки на апикальной поверхности имеют по одной ресничке, способствующей продвижению сперматозоидов. Под базальной мембраной эпителия располагается рыхлая соединительная ткань, окружающая трубочки rete testis снаружи. Генеративная функция (сперматогенез, spermatogenesis) осуществляется в извитых семенных канальцах, а эндокринная (синтез мужских половых гормонов) — в интерстициальных эндокринных клетках.