Файл: 1. Современные представления о строении и функции мембран.docx
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 10.04.2024
Просмотров: 227
Скачиваний: 0
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.

1. Современные представления о строении и функции мембран
Цитоплазматическая мембрана состоит из билипидного слоя, с погруженного в него белками. Среди липидов мембраны преобладают глицерин и сфингозамещенные липиды ( большая часть - фосфолипиды)
Гидрофильная голова обращена наружу, гидрофильные концы ( обращены внутрь) Преобладание липидов в мембрана определяет ее главные свойства:
-
гидрофобность -
текучесть
о Подвижность головы относительно хвостов
о Вращение молекулы вокруг о собственной оси
о Боковое движение соседних молекул
Белки цитоплазматической мембраны бывают двух видов:
-
Интегральные белки - химически связаны с липидами мембраны
о Ковалентные связи
о Участки гидрофобных аминокислот
-
Некоторые полностью пересекают билипидный слой ( трансмембранные белки) -
Их можно выделить только после разрушения липидного слоя
Функции:
-
Рецепторы -
Транспортные системы -
Ферменты -
Межклеточные контакты
Периферические белки - не имеют связи с липидами. Их можно отмыть с помощью буферных растворов или комплексообразующими соединениями ( ЭДТА)
Функции:
-
Цитоскелет (спектрин) -
Связь с внеклеточным матриксом ( фибронектин)
Общие функции цитоплазматической мембраны:
-
Граница клетки -
Транспорт веществ в клетку и из клетки -
Место, где протекает большинство биохимических реакций ( белки- ферменты) -
Рецепция и сигнализация: воспринимает сигналы извне и преобразует их в команды, регулирующие внутриклеточные процессы.
2. Типы ионных каналов мембраны и способы воздействия на их состояние
Виды ионных каналов: селективные и неселективные. Определяется соотношением:
• Размер поры/иона и его гидратной оболочки
• Заряд иона/канала
Селективные ионные каналы: K+, Cl", Ca2+ и т.д.
Функциональная классификация:
Управляемые ( воротные) и неуправляемые ( каналы утечки, каналы покоя)
Неуправляемые каналы открываются в покое без внешнего воздействия.
Управляемые каналы в покое закрыты и открываются при воздействии.
Основа открытия и закрытия ворот - изменение конформации белка.
Потенциал-зависимые каналы: канал открывается и закрывается при изменении заряда мембраны (мембранного потенциала). В структуре есть сенсор напряжения (заряженная белковая часть, при изменении напряжения меняет свою конформацию) .
Лиганд-зависимые ионные каналы: каналы открываются при присоединении сигнальной молекулы (лиганда). Могут активироваться как снаружи (гормоны, медиаторы), изнутри (вторичные посредники). Содержат рецептор, обладающий высоким сродством к лиганду.
Механо-чувствительные ионные каналы: открываются при растяжении мембраны. Они характерны для цитоплазматической мембраны гладких миоцитов.
Ионные каналы - мишень для действия токсинов и лекарств (местные анестетики).
Потенциал-зависимые na- каналы.
- Тетродотоксин (иглобрюх)
- Местные анестетики, лидокаин
Потенциал зависимые К-каналы:
- Яд черной мамбы
Патология хлорных каналов – муковисцидоз. Патология Na – каналов - эпилепсия с приступами лихорадки.
3. Транспорт веществ через цитоплазматическую мембрану: виды и механизмы.
Движущая сила - градиент ( разность концентраций)
Градиент концентрации - некий запас потенциальной энергии, который реализуется при движении вещества из области с высокой концентрацией в область с низкой концентрацией.
Виды трансмембранного транспорта:
Пассивный транспорт:
-
По grad с -
Без дополнительных затрат -
Виды:
о Диффузия (закон диффузии Фика – отражает скорость диффузии вещества через мембрану в зависимости от разности концентраций вещества по обе её стороны) Облегченная диффузия - пассивный транспорт при участии специальных белков- ферментов.
о Осмос
Активный транспорт:
-
Против grad c -
С доп. Затратами -
Виды
о Первично активный транспорт ( АТФ)
о Вторично активный транспорт ( grad с другого вещества)
Осмос - движение молекул растворителя через полупроницаемую мембрану из области с меньшей концентрацией растворенного вещества в область с большей концентрацией.
Осмотическое давление - величина гидростатического давления, которое нужно приложить, чтобы воспрепятствовать осмотическому движению молекул растворителя.

Осмолярность - величина осмотически активных веществ.
Изотонический раствор - раствор, осмотическое давление которого равно осмотическому давлению данного раствора. ( для клеток человека - рр NaCl 0,9%)
Очаг воспаления сопровождается отеком. Крупные молекулы разрушаются на более мелкие. Увеличивается осмолярность. Привлекается жидкость. На этом принципе основывается применение гипертонических соляных повязок. На этом же основании применяется солевое слабительное.
Активный транспорт.
Вещество перемещается против grad C.
Первично активный транспорт:
-
Энергия гидролиза АТФ ( транспортная система - катализатор гидролиза)
о Na/K-АТФаза
о Ca-АТФаза
-
Перемещается одно ( унипорт) или несколько веществ ( контрапорт)
Вторично активный транспорт:
-
Энергия, запасенная в grad C другого вещества ( как правило натрия) -
Всегда котранспорт ( Na - пассивно, др вещество - активно)
о Однонаправленный ( симпорт) - всасывание глюкозы
о Разнонаправленный ( антипорт) - обменники ( Na/Ca обменник)
Мембрана гидрофобна - свободно проходят вещества, растворимые в липидах:
-
Стероидные гормоны ( производные холестерина) -
Недиссоциированные жирные кислоты -
Неполярные молекулы ( O2, NO, CO2, CO)
Большинство органических молекул содержат ионизированный остаток фосфорной кислоты - не проходят через мембрану и остаются снаружи/внутри.
Мелкие ионизированные и полярные молекулы перемещаются с помощью гидрофильных белковых структур: каналов и переносчиков.
Вода - полярная молекула. Но она свободно проходит через мембрану:
-
Мелкие размеры молекулы -
Подвижность липидов -
Аквапорины - водные каналы мембраны -
4. Мембранный потенциал покоя: определение, ионные механизмы его формирования.
1. Цитоплазма клетки с внутренней стороны мембраны заряжена отрицательно, благодаря анионам (неорганическим и органическим), которые не могут выходить из клетки, а с наружной стороны заряжена положительно, благодаря катионам.
2. Неорганический катион К+ свободно проходит через клеточную мембрану в покое. Он движется из области высокой концентрации (изнутри клетки) в область низкой концентрации (на наружную поверхность мембраны клетки) по ионным каналам. Это «химическая» составляющая заряда на мембране.
3. Остающиеся внутри клетки анионы и скапливающиеся снаружи клеточной мембраны катионы создают электрическое поле, и ионы К+ начинают двигаться в этом поле. Анионы внутри клетки притягивают положительные ионы К+ («электрическая» составляющая), а по концентрационному градиенту ионы К+ стремятся выйти из клетки. По своему характеру проницаемость ионов калия через мембрану нейрона носит диффузионный характер (ток утечки ионов калия) и потен¬циал покоя определяется, в основном, этим ионом.
4. В тот момент, когда влияние электрического поля будет скомпенсировано диффузионным давлением (обусловленным разностью концентраций), возникает электро-химическое равновесие. В точке равновесия ионный ток К+ наружу (выходящий ток) и внутрь клетки (входящий ток) равны.
5. Происходит разделение зарядов на поверхности мембраны – внутренняя поверхность становится более отрицательной по отношению к внешней.
6. В момент равновесия внутри клетки можно зарегистрировать отрицательный заряд, равный -90 мВ. Эта разность потенциалов называется равновесным потенциалом для К+ (Ек), который можно определить с помощью уравнения Нернста.
7. Потенциал на мембране или мембранный потенциал покоя в действительности немного меньше Ек (обычно от -65 мВ до – 80 мВ) вследствие того, что какая-то часть ионов Na+ может входить в клетку в покое.
8. Концентрация ионов Na+ в покое снаружи клетки больше, чем внутри, а внутренняя поверхность клетки заряжена отрицательно. Отрицательный заряд притягивает ионы Na+.
9. В отсутствии возбуждения небольшой выходящий ток К+ сопровождается небольшим входящим током Na+. Входящий ток ионов Na+ невелик, так как в покое клеточная мембрана почти непроницаема для ионов Na+.
10. Концентрационный градиент для ионов Na+ и К+ и, соответственно, мембранный потенциал покоя, поддерживается работой Na+/К+насоса, который обеспечивает одновременный выход 3-х ионов Na+ из клетки и 2-х ионов К+ калия внутрь клетки. Для работы Na+/К+насоса необходима энергия АТФ (активный транспорт).
11. Работа Na+/К+насоса дает свой отдельный вклад в мембранный потенциал, так как выводит из клетки больше ионов Na+, чем вносит ионов К+. Благодаря работе насоса внутренняя поверхность клетка становится еще более отрицательной, поэтому такой насос назвали электрогенным насосом.
5. Потенциал действия: определение, фазы, свойства и ионные механизмы его формирования.
Потенциал действия - быстрое изменение мембранного потенциала возбудимой клетки при действии на нее деполяризующего раздражителя как минимум пороговой величины. Причина - быстрое пассивное движение ионов через потенциалзависимые каналы мембраны. (лекция)
Потенциал действия возбудимых клеток представляет собой кратковременное фазное изменение знака потенциала на внутренней поверхности мембраны и проницаемости мембраны для ионов, вызванное внешним раздражением мембраны возбудимых клеток. Потенциал действия состоит из фаз деполяризации, реполяризации и следовых потенциалов.
Открытый ионный Инактивированный
Р
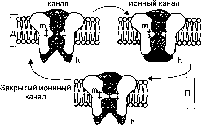 ис. 2.9. Натриевый воротный ионный канал. При деполяриза¬ции мембраны канал находится в открытом или инактивированном состояниях (закрыты h-ворота). При поляризованной мембране, т. е. когда внутренняя поверхность мембраны имеет отрицательный за¬ряд относительно наружной, ион¬ный канал находится в закрытом состоянии (m-ворота закрыты).
ис. 2.9. Натриевый воротный ионный канал. При деполяриза¬ции мембраны канал находится в открытом или инактивированном состояниях (закрыты h-ворота). При поляризованной мембране, т. е. когда внутренняя поверхность мембраны имеет отрицательный за¬ряд относительно наружной, ион¬ный канал находится в закрытом состоянии (m-ворота закрыты).Д — деполяризация мембраны, П — поляризация мембраны в состоянии покоя.
Фаза деполяризации возникает в результате открытия в мембране потенциалзависимых натриевых ионных каналов (рис.2.9). При этом коэффициент проницаемости мембраны для ионов Na+ возрастает в сотни раз, т. е. с PNa+ = 0,04 до PNa+ = 20. Потенциал действия возникает по закону «все или ничего». Согласно этому закону величина потенциа¬ла действия остается посто¬янной, независимо от силы вызвавшего его стимула. Стимулом для генерации потенциала действия явля¬ется быстрая кратковре¬менная деполяризация мембраны возбудимой клетки, которая называется критическим уровнемдеполяризащи (Ек). Величина потенциала на мембра¬не (Ет), необходимая для достижения критического уровня деполяриза¬ции, называется порсговым потенциалом (Ет—Ек), или порсгом генерации потенциала действия (рис. 2.10). Амплитуда критического уровня деполя- ризациии мембраны нервного волокна и мышечной клетки в среднем меньше величины потенциала покоя на 15—20 мВ. Пороговый сдвиг мем¬бранного потенциала возникает лишь в том случае, если натриевый ток (ISa), направленный внутрь клетки начинает превышать противоположно направленный калиевый ток (1К). £сли уменьшение мембранного потен¬циала происходит медленно в течение нескольких миллисекунд, то инак- тивационные ворота в натриевых ионных каналах инактивируются раньше, чем произойдет открывание достаточного количества активационных во¬рот. В результате натриевый ток (/^ ) будет недостаточным для открытия потенциалзависимых натриевых каналов. Подобное явление называется аккомодацией мембраны В этом случае не будет возникать потенциал дей¬ствия. Деполяризация мембраны начинается с активации инактивирован¬ных потенциалзависимых натриевых ионных каналов, и только после это¬го натриевые каналы открываются. Ионы т + через открытые потенциалзависимые натриевые каналы устремляются по концентрационному гради¬енту внутрь клетки (натриевый ток INa), заряжая внутреннюю поверхность мембраны положительным зарядом. При этом отношение коэффициентов проницаемостей мембраны для ионов калия и натрия составляет иную ве¬личину относительно состояния покоя: Рк : PNa = 1 : 20, а генерация по¬тенциала действия обусловлена высокой проницаемостью мембраны для ионов Na+. Деполяризация мембраны клетки увеличивает число открытых потенциалзависимых натриевых каналов, что существенно повышает на¬триевую проницаемость. В результате потенциал на мембране возбудимой клетки за 1—2 мс стремится достичь величины натриевого равновесного потенциала. При этом потенциал на внутренней поверхности мембраны меняет знак и его величина достигает +40 мВ (явление реверсии заряда мембраны).