Файл: 1. Современные представления о строении и функции мембран.docx
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 10.04.2024
Просмотров: 232
Скачиваний: 0
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.

Реполяризация. Во время реверсии заряда мембраны величина потенциала Нернста для ионов натрия приближается к их равновесному потенциалу, что вызывает через 1—2 мс после начала деполяризации инактивацию потенциалзависимых натриевых ионных каналов. При этом инактивацион- ные ворота натриевых ионных каналов не открываются до тех пор, пока мембранный потенциал покоя не восстанавливается до своего исходного уровня. В результате прекращается входящий натриевый ток. Одновременно под влиянием деполяризации мембраны открываются потенциалзависимые калиевые ионные каналы и увеличивается выходящий калиевый ток, который стремится сместить мембранный потенциал в сторону калиевого равновесного потенциала и, таким образом, реполяризует мембрану (рис, 2.11). Мембранный потенциал приближается к величине потенциала покоя, но в мембране еще остаются открытыми калиевые ионные каналы и инактивироваными — натриевые ионные каналы.
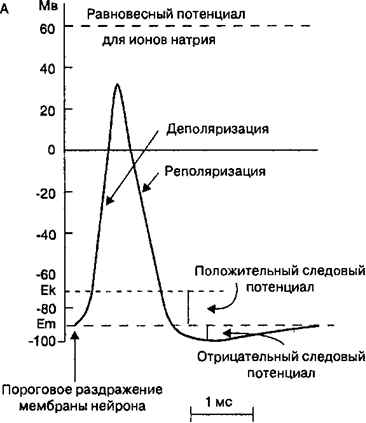 Рис. 2.10. Потенциал дейст¬вия. Изменения мембранного потен¬циала (А), которые называются потенциалом действия нервов и мышц, являются результатом из¬менения ионной проницаемости мембраны для Na’ и К* (Б). В за¬висимости от иона, участвующего в генерации потенциала действия, изменение ионной проницаемости мембраны происходит в результате открытия, закрытия или инакти- вирования ионных каналов. Боль¬шинство потенциалзависимых на¬триевых ионных каналов открыва¬ется, когда деполяризация дости¬гает порогового уровня (Ек). По¬сле спонтанной инактивации этих каналов открываются калиевые ионные каналы, что вызывает ре¬поляризацию мембраны. В пери-од, когда натриевые ионные кана¬лы находятся в инактивированном состоянии, мембрана клетки явля¬ется рефрактерной. Натриевые ионные каналы функционируют примерно в 10 раз быстрее, чем калиевые.
Рис. 2.10. Потенциал дейст¬вия. Изменения мембранного потен¬циала (А), которые называются потенциалом действия нервов и мышц, являются результатом из¬менения ионной проницаемости мембраны для Na’ и К* (Б). В за¬висимости от иона, участвующего в генерации потенциала действия, изменение ионной проницаемости мембраны происходит в результате открытия, закрытия или инакти- вирования ионных каналов. Боль¬шинство потенциалзависимых на¬триевых ионных каналов открыва¬ется, когда деполяризация дости¬гает порогового уровня (Ек). По¬сле спонтанной инактивации этих каналов открываются калиевые ионные каналы, что вызывает ре¬поляризацию мембраны. В пери-од, когда натриевые ионные кана¬лы находятся в инактивированном состоянии, мембрана клетки явля¬ется рефрактерной. Натриевые ионные каналы функционируют примерно в 10 раз быстрее, чем калиевые.Следовые потенциалы. В возбудимых клетках фаза реполяризации переходит в следовые потенциалы, ко-торые подразделяют на следовую деполяризацию и следовую гиперполяриза¬цию. Следовые потенциа¬лы обусловлены тем, что натриевые и калиевые ионные каналы не возвра¬тились к своему исходному состоянию после быстрых фаз деполяризации и репо¬ляризации. При этом чем медленнее в фазу деполя¬ризации открываются по-тенциалзависимые калие¬вые ионные каналы, тем длительнее будет следовая гиперполяризация и тем медленнее в мембране клетки будет восстанавли¬
ваться ее нормальная возбудимость. Величина потенциала покоя мембра¬ны нейронов составляет —60 мВ, а мембраны нервного волокна и скелет¬ной мышечной клетки —90мВ, что свидетельствует о более высокой возбу¬димости сомы нейронов, чем их аксонов и, тем более, скелетных мышеч¬ных волокон.
При этом амплитуда потенциала действия в нервных волокнах состав¬ляет 110 мВ, что на +20 мВ превышает величину потенциала покоя. В ске¬летных мышечных клетках во время фазы деполяризации величина потен¬циала на мембране достигает максимального значения +30—40 мВ. Поэто¬му амплитуда потенциала действия в клетках скелетных мышц составляет 120—130 мВ. Эти различия в амплитуде реверсии заряда мембраны нерв¬ных и мышечных волокон обусловливают большую скорость протекания возбуждения в нервных волокнах, а следовательно, более высокую их ла¬бильность по сравнению с мышечными волокнами.
6. Локальный ответ: определение, свойства, отличия от ПД.
При действии электрического стимула подпороговой силы уменьшение мембранного потенциала оказывается недостаточным для того, чтобы натриевый ток (INJ стал больше, чем сумма калиевого выходящего тока или тока утечки ионов калия. При этом мембранный потенциал уменьшается на несколько милливольт, но в результате реполяризации восстанавливается до величины мембранного потенциала покоя. Подобные пассивные или электротонические изменения потенциала на мембране, которые не достигают критического уровня деполяризации под действием деполяризующего электрического тока подпороговой величины, называются локальным ответом. Для локального ответа характерны следующие физиологические проявления на мембране нервной клетки или нервного волокна: амплитуда локального ответа не имеет линейной зависимости от силы подпорогового электрического раздражения; отсутствует проявление закона «все или ничего»; в период локального ответа мембрана клетки или нервного волокна не только сохраняет, но и имеет повышенную (супернор- мальную) возбудимость. Следовательно, локальный ответ не имеет четкого порога возникновения, а в период его развития отсутствует абсолютная рефрактерность нервного волокна. Во время локального ответа мембрана нервной клетки имеет высокую способность к регенеративному увеличению

ионной натриевой проницаемости, поскольку амплитуда мембранного потенциала снижается электротонически. Локальные электротонические изменения потенциала на мембране нервного волокна после прекращения действия кратковременного импульса электрического тока подвергаются обратному самостоятельному развитию. При действии на возбудимую мембрану серией подпороговых электрических стимулов происходит суммирование амплитуды локального ответа. Если амплитуда локального ответа достигает порога возбудимости мембраны, то потенциалзависимые натриевые ионные каналы открываются по закону «все или ничего» и происходит генерация потенциала действия. На рис. 2.12 показано суммирование амплитуды локального ответа по типу деполяризации при действии на мембрану нервного волокна серией подпороговых электрических стимулов. Локальный ответ нервного волокна всегда сопровождается повышением возбудимости мембраны клеток.
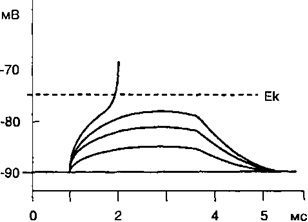
Рис. 2.12. Влияние подпорогового элек¬трического раздражения возбудимой мем¬браны на степень ее деполяризации. При раздражении мембраны возбудимых кле¬ток электрическим током подпороговой силы происходит электротоническое уменьшение потенциала на мембране и/или активация по-тенциалзависимых натриевых каналов. Про¬цесс является обратимым, и мембранный по¬тенциал возвращается к исходному уровню, не достигая критического уровня деполяризации (Ек). В период локальных изменений мембран¬ного потенциала мембрана возбудимых клеток имеет супернормальную возбудимость.
7. Изменение возбудимости во время генерации потенциала действия. Понятие о рефрактерности, причины её возникновения.
В фазу деполяризации потенциала действия потенциалзависимые натриевые ионные каналы кратковременно открываются, но затем инактивируются h-ворота (см. рис. 2.9). В период инактивации натриевых ионных каналов возбудимые клетки не способны реагировать повышением натриевой проницаемости на повторный стимул. Поэтому во время фазы деполяризации мембрана не может генерировать потенциал действия в ответ на действие пороговых или сверхпороговых раздражителей. Это состояние называется абсолютной рефрактерностъю> время которой составляет в нервных волокнах 0,5—1,0 мс, а в скелетных мышечных клетках —в среднем 2 мс. Период абсолютной рефрактерности заканчивается после того, как снижается количество инактивированных натриевых каналов и постепенно увеличивается количество натриевых каналов, находящихся в закрытом состоянии. Происходят эти процессы во время фазы реполяризации, когда уменьшению количества потенциалзависимых натриевых ионных каналов, находящихся в состоянии инактивации, соответствует период относительной рефрактерности. Период относительной рефрактерности характеризуется тем, что лишь некоторая часть потенциалзависимых натриевых ионных каналов переходит в закрытое состояние, а в силу этого порог возбудимости мембраны клетки имеет более высокие значения, чем в исходном состоянии. Поэтому возбудимые клетки в период относительной рефрактерности могут генерировать потенциалы действия, но при воздействии на них раздражителей сверхпороговой силы. Однако из-за небольшого количества потенциалзависимых натриевых ионных каналов, находящихся в закрытом состоянии, амплитуда генерируемых при этом потенциалов действия будет меньше, чем в условиях исходной возбудимости нервной или мышечной клетки.
В клетках возбудимых тканей максимальное количество генерируемых потенциалов действия в единицу времени обусловлено двумя факторами: длительностью потенциала действия и длительностью периода абсолютной рефрактерности после каждого импульса. На этом основании в физиологии формулируется современное понятие
лабильности: чем меньше период абсолютной рефрактерности при возбуждении возбудимой ткани, тем выше ее функциональная подвижность или лабильность, тем больше в ней генерируется потенциалов действия в единицу времени.
При непрерывной стимуляции нерва электрическим током лабильность нерва зависит от частоты и силы раздражения. В зависимости от частоты и силы раздражения нерва сокращение иннервируемой им мышцы может быть максимальной или минимальной амплитуды. Эти явления были названы соответственно оптимумом и пессимумом (Н. Е. Введенский). Максимальное (оптимально большое) сокращение мышцы возникает в том случае, если каждый последующий электрический стимул действует на нерв в периоде его состояния супернормальной возбудимости после предыдущего потенциала действия. Минимальное (или пессимальное) сокращение мышцы возникает в том случае, если каждый последующий электрический стимул действует на нерв, находящийся в периоде относительной рефрактерности после предыдущего потенциала действия. Поэтому значения оптимальной частоты раздражения нерва всегда меньше, чем значения пессимальной частоты раздражения.
8. Общие свойства возбудимых тканей. Возбудимость, критерии её оценки.
Свойства:
1) Возбудимость – это способность отвечать возбуждением (генерацией потенциала действия) в ответ на действие раздражителей любой природы.
2) проводимость – способность ткани передавать возникшее возбуждение за счет электрического сигнала от места раздражения по длине возбудимой ткани;
3) Рефрактерность – временное снижение возбудимости одновременно с возникшим в ткани возбуждением. Рефрактерность бывает абсолютной (нет ответа ни на какой раздражитель) и относительной (возбудимость восстанавливается, и ткань отвечает на подпороговый или сверхпороговый раздражитель);
4) лабильность (функциональная подвижность) - способность ткани воспроизводить определенное число волн возбуждения в единицу времени в точном соответствии с ритмом наносимых раздражений. Это свойство характеризует скорость возникновения возбуждения. Показатель лабильности: максимальное количество волн возбуждения в данной ткани: нервные волокна - 500-1000 импульсов в секунду, мышечная ткань - 200-250 импульсов в секунду, синапс - 100-125 импульсов в секунду. Лабильность зависит от уровня обменных процессов в ткани, возбудимости, рефрактерности.