Файл: Глоссарий Клетка это структурная единица организма. Гликокаликс.docx
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 26.04.2024
Просмотров: 14
Скачиваний: 0
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.

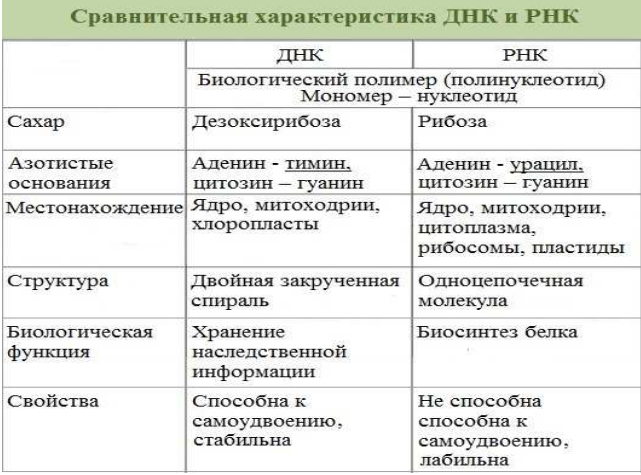
Строение, функции и классификация нуклеиновых кислот. Строение ДНК.
Нуклеиновые кислоты – материальный субстрат наследственности и изменчивости, это макромолекулы, биополимеры мономерами которых являются нуклеотиды. Непрерывность существования живой клетки обусловлена двумя фундаментальными свойствами жизни:
наследственностью и изменчивостью, которые обеспечиваются нуклеиновыми кислотами. Впервые нуклеиновые кислоты были открыты в 1868 г. (Мишер).
Нуклеиновые кислоты – это высокомолекулярные соединения с определенной последовательностью мономеров - нуклеотидов. Информация о структуре белка, хранится и воспроизводится при помощи нуклеиновых кислот.
Типы нуклеиновых кислот:
1. Дезоксирибонуклеиноваякислота (ДНК)
2. Рибонуклеиноваякислота (РНК)
Важнейшим процессом, происходящим во всех клетках, является синтез
белка. Информация о последовательности аминокислот, составляющих первичную
структуру белка, заключена в ДНК. Молекула дезоксирибонуклеиновой кислоты
(ДНК) состоит из двух спирально закрученных цепей. Общепринятая модель
структуры ДНК была впервые предложена Уотсон и Криком в 1953г. Модель
строения основывалась на следующих данных:
1. ДНК представляет собой полимер, состоящий из нуклеотидов, соединенных 3/ - 5/
фосфодиэфирными связями.
2. Состав нуклеотидов в ДНК подчиняется правилам Чаргаффа(правило
комплементарности). В молекуле ДНК количество аденина равно количеству
тимина, а количество гуанина — количеству цитозина: А = Т, Г = Ц.
3. Рентгенограммы волокон ДНК, впервые полученные Уилкинсом и Франклин указывают на то, что молекулы обладают спиральной структурой.
4. Структура нативной ДНК стабилизируется водородными связями.
8) Особенности пространственной организации ДНК. Свойства и функции ДНК.
Первичная структура:
• Мономером ДНК является нуклеотид ДНК. Нуклеотиды соединяются между собой фосфодиэфирной связью, образуя полинуклеотидную цепь. На полинуклеотидной цепи ДНК различают два конца 5’ и 3’, начало и конец. Образование фосфодиэфирной связи обеспечивает фермент ДНК-полимераза. Благодаря ДНК-полимеразе происходит наращивание полинуклеотидной цепи только на одном конце (3’). Начало любой полинуклеотидной цепи всегда несет фосфатную группу в положении 5’.
• Полинуклеотидная цепь – это первичная структура ДНК.
Вторичная:
Вторичная структура - это две антипараллельные комплементарные полинуклеотидные цепи, соединенные между собой водородными связями, и образующие спираль. Диаметр спирали
составляет 2 нм, длина шага- 3.4 нм, в каждый виток входит 10 пар нуклеотидов. Комплементарность в молекуле ДНК: Аденин – Тимин, Гуанин – Цитозин.
• Аденин одной цепи соединяется двумя водородными связями с тимином другой цепи. Между гуанином и цитозином – три водородные связи.
Такое соединение азотистых оснований обеспечивает прочную связь двух цепей.
• Антипараллельность в молекуле ДНК: 5 конец одной цепи соединяется с 3-концом другой.
Третичная:
Третичная структура – упакованная молекула ДНК. Двойная спираль ДНК на некоторых участках может подвергаться дальнейшей спирализации. Супер спиральная структура обеспечивает экономную упаковку огромной молекулы ДНК. Упаковка происходит при помощи гистоновых и негистоновых белков. Одна молекула ДНК в комплексе с белками в ядрах образует нить хроматина. При делении хроматин конденсируется.
Функции ДНК:
1. хранение наследственной информации;
2. передача наследственного материала;
3. реализация наследственной информации в процессе синтеза белка
9) Особенности строения митохондриальной ДНК.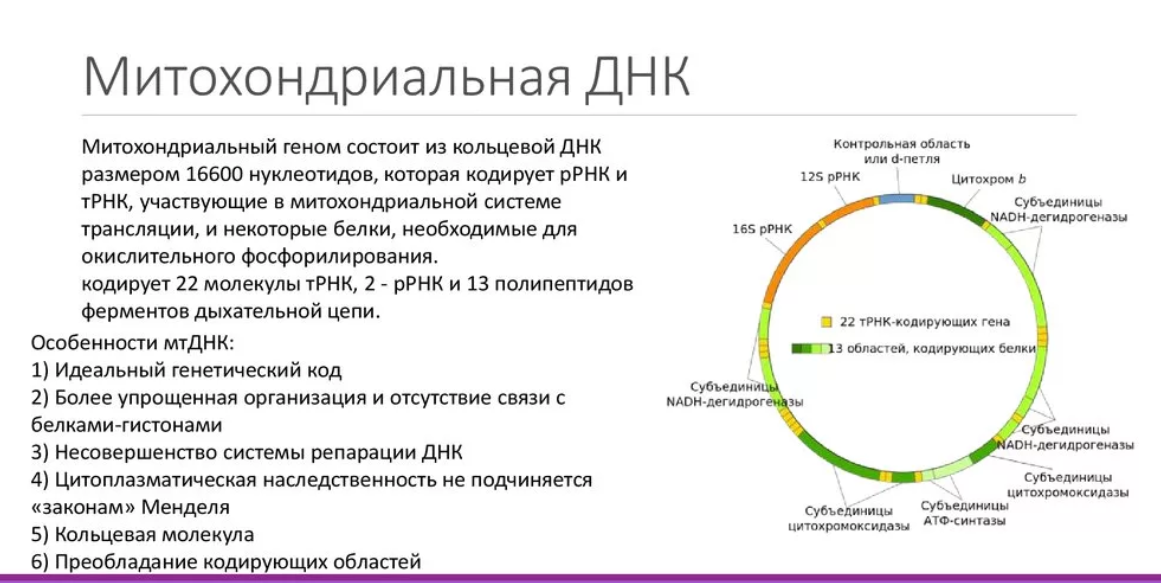
10) Репликация ДНК. Образование репликативного комплекса. Фазы репликации.
Репликация – способность к самокопированию, это одно из основных свойств наследственного материала. Репликация ДНК обеспечивает воспроизведение наследственной информации при образовании новых клеток. Клеточное деление бывает двух типов: митоз (количество хромосом не изменяется) и мейоз (количество хромосом уменьшается в два раза).
• Репликация ДНК происходит полуконсервативным способом, когда каждая дочерняя молекула ДНК содержит одну материнскую и одну синтезированную цепь.
• Процесс репликации осуществляется сложным ферментным комплексом (15-20 различных белков). На каждой хромосоме работают несколько репликативных комплексов. На хромосоме имеется много точек начала репликации ДНК (ori) – в среднем около 40. Удвоение происходит непоследовательно, а одновременно во многих местах сразу.
• В каждой точке репликации начинают работать два ферментативных комплекса, двигаясь в противоположные стороны – образуются две репликативные вилки. Между вилками образуется постепенно расширяющееся «вздутие» или «глазок». Каждый комплекс реплицирует обе цепи. В конечном счете соседние зоны репликации (глазки) сливаются и вся молекула ДНК оказывается удвоенной.
Ферментативный комплекс функционирует так, что одна из двух синтезируемых им цепей растет с некоторым опережением по сравнению с другой цепью. Первая цепь называется лидирующей вторая запаздывающей. Запаздывающая же цепь образуется в виде серии
относительно коротких фрагментов – примерно по 1500 нуклеотидов (фрагменты Оказаки).
• Репликация – сложнейшей биохимический процесс, в котором принимают
участие множество белков (ферментов), обеспечивающих точность воспроизведения генетической информации, устранение и замену неправильных или поврежденных нуклеотидов (репарация ДНК) и рекомбинацию.
Характеристика репликативного комплекса.
В процессе репликации ДНК участвует сложный ферментный комплекс, включающий 15–20 белков-ферментов. Выделяют белки, подготавливающие родительскую ДНК к репликации, ферменты полимеризации, ферменты завершающие репликацию ДНК. Для осуществления репликации цепи материнской ДНК должны быть отделены друг от друга, чтобы стать матрицами, на которых будут синтезироваться комплементарные цепи дочерних молекул. Репликация ДНК состоит из следующих периодов:
а) инициация репликации(самая начальная фаза);
б) полимеризация;
в) терминация.
Репликация происходит двумя механизмами:
(1) Непрерывный синтез: ДНК-полимераза добавляет нуклеотиды к 3’ концу лидирующей
нити.
(2) Прерывистый синтез: Праймаза добавляет короткий РНК праймер впереди 5’ конца отстающей цепи. Затем ДНК-полимераза добавляет нуклеотиды к праймеру пока пробел не заполняется. ДНК полимераза I заменяет праймер на нуклеотиды ДНК, и ДНК-лигаза
сшивает короткие сегменты нуклеотидов к отстающей нити.
11) Этапы репликации ДНК. Особенности синтеза лидирующей и отстающей цепи.
Этапы репликации
1. Инициация репликации осуществляется в особых участках ДНК, обозначаемых ori (от англ. оrigin - начало). Точки начала репликации на молекуле ДНК имеют специфическую последовательность основании, богатую парами А-Т. Процесс начинается с того, что с каждой такой последовательностью связывается несколько молекул узнающих белков. Двойная спираль ДНК в этих локусах, под действием фермента геликазы, разделяется на две цепи, при этом, как правило, по обе стороны от точки начала репликации образуются области расхождения полинуклеотидных цепей – репликационные вилки, которые движутся в
противоположных от локуса ori направлениях.
• Фермент геликаза разделяет нити двойной цепи ДНК. Образующиеся при этом одинарные цепи ДНК связываются специальными дестабилизирующими белками (SSB-белки), которые растягивают остовы цепей, делая их азотистые основания доступными для связывания с комплементарными нуклеотидами. Разделение спирально закрученных цепей родительской ДНК ферментом геликазы вызывает появление супервитков перед репликационной вилкой. Фермент топоизомераза I разрывает одну цепь ДНК и дает возможность вращаться другой цепи, тем самым ослабляет напряжение в двойной спирали ДНК.
2. Полимеризация цепи ДНК
• Синтез цепи ДНК (полимеризацию нуклеотидной цепи)
Особенностью ДНК-полимеразы является ее не способность начать синтез новой поленуклеотидной цепи путем простого связывания двух нуклеозиттрифосфатов необходим 3’-ОН-конец какой-либо полинуклеотидной цепи, спаренной с матричной цепью ДНК, к которой ДНК-полимераза может лишь добавлять новые нуклеотиды. Такую полинуклеотидную цепь называют затравкой или праймером. Роль затравки для синтеза полинуклеотидных цепей ДНК в ходе репликации выполняют короткие последовательности РНК, образуемые при участии фермента РНК-праймазы.
• Специальный белок выполняет функции активатора праймазы. После чего праймаза, используя в качестве матрицы соответствующий участок однуцепочечную ДНК, синтезирует короткую РНК-затравку, или праймер. Указанная особенность ДНК-полимеразы означает, что матрицей при репликации может служить лишь цепь ДНК, несущая спаренную с ней затравку, которая имеет свободный 3’-ОН-конец. Этот фермент образует межнуклеотидную (фосфодиэфирную) связь. Для начала синтеза новой цепи для ДНК-полимеразы необходимо наличие свободного 3’-OH конца, к которому будет присоединен 5’-конец следующего нуклеотида, т.е. необходимо наличие праймера (затравки) комплементарного одной из материнских цепей. Для этого в точке начала репликации фермент РНК-праймаза синтезирует короткую цепь РНК (РНК-затравка, или праймер).
• У эукариот известно 5 разных ДНК-полимераз. Из них β и ε полимеразы участвую в репарации ДНК, γ-полимераза - в репликации митохондриальной ДНК, а α- и δ-полимеразы в
репликации ядерной РНК. Аналогом бактериальной ДНКполимеразы III у эукариот является комплекс α- и δ-ДНКполимераз (синтез цепи ДНК).
Функцию ДНК-полимеразы I выполняют: специальная нуклеаза (удаление РНК-затравки) и β-ДНКполимераза (достраивание брешей).
Особенности синтеза лидирующей и отстающей цепей.
• Поскольку синтез цепи ДНК возможен только в направлении 5’- 3’, дочерние цепи ДНК в репликативной вилке синтезируются не с одинаковой скоростью. Ферментный комплекс
периодически прекращает образование лидиру.
• Сшивание соседних фрагментов ДНК осуществляет ДНКлигаза. Этот фермент образует межнуклеотидную фосфодиэфирную связь. Молекула ДНК окажется реплицированной не полностью, если не произойдет специальный процесс репликации ее концов, или теломерных участков. В этом процессе ключевую роль играет фермент теломераза.
• Репликация заканчивается в участке гена (молекулы ДНК), содержащим несколько специфических последовательностей нуклеотидов, называемых терминационными сайтами. Это короткие (около 23 пар нуклеотидов) последовательности.
Терминация - конечный этап – завершение репликации.
• У эукариот проблема недорепликации – концов линейных молекул ДНК мешается с помощью специального фермента – теломеразы, которая перед концом репликации наращивает недорепликированные 5‘ – концы ДНК дочерних рибосом.
• Плотность упаковки теломер позволяет отнести их к гетерохромативновой структуре хромосом.
12) Особенности репликации теломерной ДНК. Роль теломер и теломераз в репликации концевых отделов ДНК.
Теломеры – концевые участки
Нуклеиновые кислоты – материальный субстрат наследственности и изменчивости, это макромолекулы, биополимеры мономерами которых являются нуклеотиды. Непрерывность существования живой клетки обусловлена двумя фундаментальными свойствами жизни:
наследственностью и изменчивостью, которые обеспечиваются нуклеиновыми кислотами. Впервые нуклеиновые кислоты были открыты в 1868 г. (Мишер).
Нуклеиновые кислоты – это высокомолекулярные соединения с определенной последовательностью мономеров - нуклеотидов. Информация о структуре белка, хранится и воспроизводится при помощи нуклеиновых кислот.
Типы нуклеиновых кислот:
1. Дезоксирибонуклеиноваякислота (ДНК)
2. Рибонуклеиноваякислота (РНК)
Важнейшим процессом, происходящим во всех клетках, является синтез
белка. Информация о последовательности аминокислот, составляющих первичную
структуру белка, заключена в ДНК. Молекула дезоксирибонуклеиновой кислоты
(ДНК) состоит из двух спирально закрученных цепей. Общепринятая модель
структуры ДНК была впервые предложена Уотсон и Криком в 1953г. Модель
строения основывалась на следующих данных:
1. ДНК представляет собой полимер, состоящий из нуклеотидов, соединенных 3/ - 5/
фосфодиэфирными связями.
2. Состав нуклеотидов в ДНК подчиняется правилам Чаргаффа(правило
комплементарности). В молекуле ДНК количество аденина равно количеству
тимина, а количество гуанина — количеству цитозина: А = Т, Г = Ц.
3. Рентгенограммы волокон ДНК, впервые полученные Уилкинсом и Франклин указывают на то, что молекулы обладают спиральной структурой.
4. Структура нативной ДНК стабилизируется водородными связями.
8) Особенности пространственной организации ДНК. Свойства и функции ДНК.
Первичная структура:
• Мономером ДНК является нуклеотид ДНК. Нуклеотиды соединяются между собой фосфодиэфирной связью, образуя полинуклеотидную цепь. На полинуклеотидной цепи ДНК различают два конца 5’ и 3’, начало и конец. Образование фосфодиэфирной связи обеспечивает фермент ДНК-полимераза. Благодаря ДНК-полимеразе происходит наращивание полинуклеотидной цепи только на одном конце (3’). Начало любой полинуклеотидной цепи всегда несет фосфатную группу в положении 5’.
• Полинуклеотидная цепь – это первичная структура ДНК.
Вторичная:
Вторичная структура - это две антипараллельные комплементарные полинуклеотидные цепи, соединенные между собой водородными связями, и образующие спираль. Диаметр спирали
составляет 2 нм, длина шага- 3.4 нм, в каждый виток входит 10 пар нуклеотидов. Комплементарность в молекуле ДНК: Аденин – Тимин, Гуанин – Цитозин.
• Аденин одной цепи соединяется двумя водородными связями с тимином другой цепи. Между гуанином и цитозином – три водородные связи.
Такое соединение азотистых оснований обеспечивает прочную связь двух цепей.
• Антипараллельность в молекуле ДНК: 5 конец одной цепи соединяется с 3-концом другой.
Третичная:
Третичная структура – упакованная молекула ДНК. Двойная спираль ДНК на некоторых участках может подвергаться дальнейшей спирализации. Супер спиральная структура обеспечивает экономную упаковку огромной молекулы ДНК. Упаковка происходит при помощи гистоновых и негистоновых белков. Одна молекула ДНК в комплексе с белками в ядрах образует нить хроматина. При делении хроматин конденсируется.
Функции ДНК:
1. хранение наследственной информации;
2. передача наследственного материала;
3. реализация наследственной информации в процессе синтеза белка
9) Особенности строения митохондриальной ДНК.
10) Репликация ДНК. Образование репликативного комплекса. Фазы репликации.
Репликация – способность к самокопированию, это одно из основных свойств наследственного материала. Репликация ДНК обеспечивает воспроизведение наследственной информации при образовании новых клеток. Клеточное деление бывает двух типов: митоз (количество хромосом не изменяется) и мейоз (количество хромосом уменьшается в два раза).
• Репликация ДНК происходит полуконсервативным способом, когда каждая дочерняя молекула ДНК содержит одну материнскую и одну синтезированную цепь.
• Процесс репликации осуществляется сложным ферментным комплексом (15-20 различных белков). На каждой хромосоме работают несколько репликативных комплексов. На хромосоме имеется много точек начала репликации ДНК (ori) – в среднем около 40. Удвоение происходит непоследовательно, а одновременно во многих местах сразу.
• В каждой точке репликации начинают работать два ферментативных комплекса, двигаясь в противоположные стороны – образуются две репликативные вилки. Между вилками образуется постепенно расширяющееся «вздутие» или «глазок». Каждый комплекс реплицирует обе цепи. В конечном счете соседние зоны репликации (глазки) сливаются и вся молекула ДНК оказывается удвоенной.
Ферментативный комплекс функционирует так, что одна из двух синтезируемых им цепей растет с некоторым опережением по сравнению с другой цепью. Первая цепь называется лидирующей вторая запаздывающей. Запаздывающая же цепь образуется в виде серии
относительно коротких фрагментов – примерно по 1500 нуклеотидов (фрагменты Оказаки).
• Репликация – сложнейшей биохимический процесс, в котором принимают
участие множество белков (ферментов), обеспечивающих точность воспроизведения генетической информации, устранение и замену неправильных или поврежденных нуклеотидов (репарация ДНК) и рекомбинацию.
Характеристика репликативного комплекса.
В процессе репликации ДНК участвует сложный ферментный комплекс, включающий 15–20 белков-ферментов. Выделяют белки, подготавливающие родительскую ДНК к репликации, ферменты полимеризации, ферменты завершающие репликацию ДНК. Для осуществления репликации цепи материнской ДНК должны быть отделены друг от друга, чтобы стать матрицами, на которых будут синтезироваться комплементарные цепи дочерних молекул. Репликация ДНК состоит из следующих периодов:
а) инициация репликации(самая начальная фаза);
б) полимеризация;
в) терминация.
Репликация происходит двумя механизмами:
(1) Непрерывный синтез: ДНК-полимераза добавляет нуклеотиды к 3’ концу лидирующей
нити.
(2) Прерывистый синтез: Праймаза добавляет короткий РНК праймер впереди 5’ конца отстающей цепи. Затем ДНК-полимераза добавляет нуклеотиды к праймеру пока пробел не заполняется. ДНК полимераза I заменяет праймер на нуклеотиды ДНК, и ДНК-лигаза
сшивает короткие сегменты нуклеотидов к отстающей нити.
11) Этапы репликации ДНК. Особенности синтеза лидирующей и отстающей цепи.
Этапы репликации
1. Инициация репликации осуществляется в особых участках ДНК, обозначаемых ori (от англ. оrigin - начало). Точки начала репликации на молекуле ДНК имеют специфическую последовательность основании, богатую парами А-Т. Процесс начинается с того, что с каждой такой последовательностью связывается несколько молекул узнающих белков. Двойная спираль ДНК в этих локусах, под действием фермента геликазы, разделяется на две цепи, при этом, как правило, по обе стороны от точки начала репликации образуются области расхождения полинуклеотидных цепей – репликационные вилки, которые движутся в
противоположных от локуса ori направлениях.
• Фермент геликаза разделяет нити двойной цепи ДНК. Образующиеся при этом одинарные цепи ДНК связываются специальными дестабилизирующими белками (SSB-белки), которые растягивают остовы цепей, делая их азотистые основания доступными для связывания с комплементарными нуклеотидами. Разделение спирально закрученных цепей родительской ДНК ферментом геликазы вызывает появление супервитков перед репликационной вилкой. Фермент топоизомераза I разрывает одну цепь ДНК и дает возможность вращаться другой цепи, тем самым ослабляет напряжение в двойной спирали ДНК.
2. Полимеризация цепи ДНК
• Синтез цепи ДНК (полимеризацию нуклеотидной цепи)
Особенностью ДНК-полимеразы является ее не способность начать синтез новой поленуклеотидной цепи путем простого связывания двух нуклеозиттрифосфатов необходим 3’-ОН-конец какой-либо полинуклеотидной цепи, спаренной с матричной цепью ДНК, к которой ДНК-полимераза может лишь добавлять новые нуклеотиды. Такую полинуклеотидную цепь называют затравкой или праймером. Роль затравки для синтеза полинуклеотидных цепей ДНК в ходе репликации выполняют короткие последовательности РНК, образуемые при участии фермента РНК-праймазы.
• Специальный белок выполняет функции активатора праймазы. После чего праймаза, используя в качестве матрицы соответствующий участок однуцепочечную ДНК, синтезирует короткую РНК-затравку, или праймер. Указанная особенность ДНК-полимеразы означает, что матрицей при репликации может служить лишь цепь ДНК, несущая спаренную с ней затравку, которая имеет свободный 3’-ОН-конец. Этот фермент образует межнуклеотидную (фосфодиэфирную) связь. Для начала синтеза новой цепи для ДНК-полимеразы необходимо наличие свободного 3’-OH конца, к которому будет присоединен 5’-конец следующего нуклеотида, т.е. необходимо наличие праймера (затравки) комплементарного одной из материнских цепей. Для этого в точке начала репликации фермент РНК-праймаза синтезирует короткую цепь РНК (РНК-затравка, или праймер).
• У эукариот известно 5 разных ДНК-полимераз. Из них β и ε полимеразы участвую в репарации ДНК, γ-полимераза - в репликации митохондриальной ДНК, а α- и δ-полимеразы в
репликации ядерной РНК. Аналогом бактериальной ДНКполимеразы III у эукариот является комплекс α- и δ-ДНКполимераз (синтез цепи ДНК).
Функцию ДНК-полимеразы I выполняют: специальная нуклеаза (удаление РНК-затравки) и β-ДНКполимераза (достраивание брешей).
Особенности синтеза лидирующей и отстающей цепей.
• Поскольку синтез цепи ДНК возможен только в направлении 5’- 3’, дочерние цепи ДНК в репликативной вилке синтезируются не с одинаковой скоростью. Ферментный комплекс
периодически прекращает образование лидиру.
• Сшивание соседних фрагментов ДНК осуществляет ДНКлигаза. Этот фермент образует межнуклеотидную фосфодиэфирную связь. Молекула ДНК окажется реплицированной не полностью, если не произойдет специальный процесс репликации ее концов, или теломерных участков. В этом процессе ключевую роль играет фермент теломераза.
• Репликация заканчивается в участке гена (молекулы ДНК), содержащим несколько специфических последовательностей нуклеотидов, называемых терминационными сайтами. Это короткие (около 23 пар нуклеотидов) последовательности.
Терминация - конечный этап – завершение репликации.
• У эукариот проблема недорепликации – концов линейных молекул ДНК мешается с помощью специального фермента – теломеразы, которая перед концом репликации наращивает недорепликированные 5‘ – концы ДНК дочерних рибосом.
| короткими, | повторяющимися | последовательностями | | | |
| нуклеотидовилителомерами. | | | | | |
| • Теломеры располагаются на концах хромосом. В состав | | | | | |
| теломер | входят | специфические | белки, | не | образующие |
• Плотность упаковки теломер позволяет отнести их к гетерохромативновой структуре хромосом.
12) Особенности репликации теломерной ДНК. Роль теломер и теломераз в репликации концевых отделов ДНК.
Теломеры – концевые участки
| хромосом, | которые | не | несут |
| генетической | информации | и | |
| защищают ДНК от расщепления нуклеазами, и предотвращают от | | | |
| слияния. | | | |
| • | В | результате | репликации |
| молекула | ДНК | окажется | |
| реплицированной не полностью, если не произойдет репликации | | | |
| теломер. | Существует | фермент, | |
| восстанавливающий | длину | | |
| теломер | – | теломераза | |
| (ДНКнуклеотидилтрансфераза). | | | |
| Теломераза есть только в | | | |
| Зародышевых клетках и клетках | | | |
| опухолей, | в | соматических | |

