ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 03.02.2024
Просмотров: 1052
Скачиваний: 0

СОДЕРЖАНИЕ
ПОПУЛЯЦИОННО-ВИДОВОЙ УРОВЕНЬ ОРГАНИЗАЦИИ ЖИЗНИ
ВИДООБРАЗОВАНИЕ В ПРИРОДЕ. ЭЛЕМЕНТАРНЫЕ ЭВОЛЮЦИОННЫЕ ФАКТОРЫ
11.5. ГЕНЕТИКО-АВТОМАТИЧЕСКИЕ ПРОЦЕССЫ (ДРЕЙФ ГЕНОВ)
11.7. НАСЛЕДСТВЕННЫЙ ПОЛИМОРФИЗМ ПРИРОДНЫХ ПОПУЛЯЦИЙ.
11.9. ПРОИСХОЖДЕНИЕ БИОЛОГИЧЕСКОЙ ЦЕЛЕСООБРАЗНОСТИ
ДЕЙСТВИЕ ЭЛЕМЕНТАРНЫХ ЭВОЛЮЦИОННЫХ ФАКТОРОВ
12.1. ПОПУЛЯЦИЯ ЛЮДЕЙ. ДЕМ, ИЗОЛЯТ
12.2. ВЛИЯНИЕ ЭЛЕМЕНТАРНЫХ ЭВОЛЮЦИОННЫХ ФАКТОРОВ НА ГЕНОФОНДЫ ЧЕЛОВЕЧЕСКИХ ПОПУЛЯЦИЙ
12.3. ГЕНЕТИЧЕСКОЕ РАЗНООБРАЗИЕ
13.1. ЭВОЛЮЦИЯ ГРУПП ОРГАНИЗМОВ
13.2. СООТНОШЕНИЕ ОНТО- И ФИЛОГЕНЕЗА
13.4. Организм как целое в историческом
Соотносительные преобразования органов
13.5. СОВРЕМЕННАЯ СИСТЕМА ОРГАНИЧЕСКОГО МИРА
ФИЛОГЕНЕЗ СИСТЕМ ОРГАНОВ ХОРДОВЫХ
14.2. ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
И ДАЛЬНЕЙШАЯ ЭВОЛЮЦИЯ ЧЕЛОВЕКА
15.3. ХАРАКТЕРИСТИКА ОСНОВНЫХ ЭТАПОВ АНТРОПОГЕНЕЗА
15.4. ВНУТРИВИДОВАЯ ДИФФЕРЕНЦИАЦИЯ ЧЕЛОВЕЧЕСТВА
БИОГЕОЦЕНОТИЧЕСКИЙ УРОВЕНЬ ОРГАНИЗАЦИИ ЖИЗНИ
16.1. БИОГЕОЦЕНОЗ - ЭЛЕМЕНТАРНАЯ ЕДИНИЦА БИОГЕОЦЕНОТИЧЕСКОГО УРОВНЯ ОРГАНИЗАЦИИ ЖИЗНИ
17.2. ЧЕЛОВЕК КАК ОБЪЕКТ ДЕЙСТВИЯ ЭКОЛОГИЧЕСКИХ ФАКТОРОВ.
АДАПТАЦИЯ ЧЕЛОВЕКА К СРЕДЕ ОБИТАНИЯ
17.4. РОЛЬ АНТРОПОГЕННЫХ ФАКТОРОВ
В ЭВОЛЮЦИИ ВИДОВ И БИОГЕОЦЕНОЗОВ
МЕДИЦИНСКАЯ ПАРАЗИТОЛОГИЯ. ОБЩИЕ ВОПРОСЫ
18.2. ФОРМЫ МЕЖВИДОВЫХ БИОТИЧЕСКИХ СВЯЗЕЙ В БИОЦЕНОЗАХ
18.3. КЛАССИФИКАЦИЯ ПАРАЗИТИЗМА
18.4. РАСПРОСТРАНЕННОСТЬ ПАРАЗИТИЗМА
18.5. ПРОИСХОЖДЕНИЕ ПАРАЗИТИЗМА
18.6. АДАПТАЦИИ К ПАРАЗИТИЧЕСКОМУ ОБРАЗУ ЖИЗНИ. ОСНОВНЫЕ ТЕНДЕНЦИИ
18.8. ФАКТОРЫ ВОСПРИИМЧИВОСТИ ХОЗЯИНА К ПАРАЗИТУ
18.9. ДЕЙСТВИЕ ХОЗЯИНА НА ПАРАЗИТА
18.10. СОПРОТИВЛЕНИЕ ПАРАЗИТОВ РЕАКЦИЯМ ИММУНИТЕТА ХОЗЯИНА
18.11. ВЗАИМООТНОШЕНИЯ В СИСТЕМЕ ПАРАЗИТ - ХОЗЯИН НА УРОВНЕ ПОПУЛЯЦИЙ
18.12. СПЕЦИФИЧНОСТЬ ПАРАЗИТОВ ПО ОТНОШЕНИЮ К ХОЗЯИНУ
18.13. ПРИРОДНО-ОЧАГОВЫЕ ЗАБОЛЕВАНИЯ
19.2. Простейшие, обитающие в полостных органах, сообщающихся с внешней средой
19.3. Простейшие, обитающие в тканях
19.4. ПРОСТЕЙШИЕ — ФАКУЛЬТАТИВНЫЕ ПАРАЗИТЫ ЧЕЛОВЕКА
20.1. ТИП ПЛОСКИЕ ЧЕРВИ PLATHELMINTHES
20.2. ТИП КРУГЛЫЕ ЧЕРВИ NEMATHELMINTHES
21.1. КЛАСС ПАУКООБРАЗНЫЕ ARACHNOIDEA
ЭВОЛЮЦИЯ ПАРАЗИТОВ И ПАРАЗИТИЗМА ПОД ДЕЙСТВИЕМ АНТРОПОГЕННЫХ ФАКТОРОВ
23.1. ПРОИСХОЖДЕНИЕ ЯДОВИТОСТИ
23.2. ЧЕЛОВЕК И ЯДОВИТЫЕ ЖИВОТНЫЕ
24.1. СОВРЕМЕННЫЕ КОНЦЕПЦИИ БИОСФЕРЫ
24.2. СТРУКТУРА И ФУНКЦИИ БИОСФЕРЫ
25.2. ПУТИ ВОЗДЕЙСТВИЯ ЧЕЛОВЕЧЕСТВА
В процессе эволюции первая характеристика многоклеточности достигается наиболее просто: среди простейших уже имеется огромное количество видов колониальных организмов. Остальные черты многоклеточности связаны с возникновением многослойности, обеспечивающей как различные условия существования клеткам, расположенным на поверхности и внутри тела, так и появление внутренней среды, объединяющей их воедино. На рис. 13.10 представлены различные варианты объединения клеток в надклеточные комплексы. Ясно, что только интеграция клеток в шаровидную структуру дает им возможность оказаться в разных условиях, дифференцироваться и взаимодействовать друг с другом.
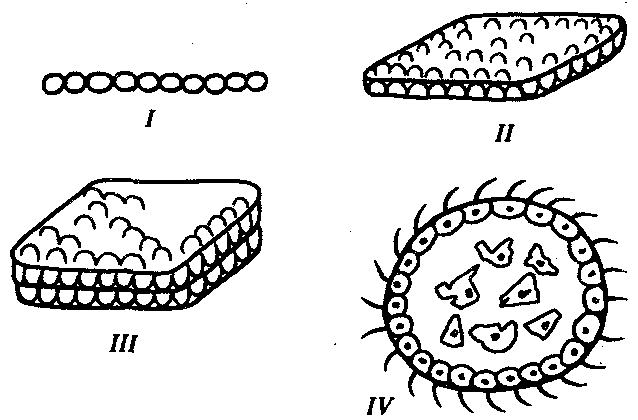
Рис. 13.10. Варианты объединения клеток в надклеточные образования:
I—нитевидная колония клеток, II—колония в виде однослойного пласта клеток, III—двуслойная колония клеток, IV—сферический многоклеточный организм с клетками разных типов
Родоначальником многоклеточных в настоящее время считают шаровидную колонию жгутиковых, половые клетки которых перемещались в глубь колонии, а соматические первично выполняли как функцию перемещения всей колонии в пространстве, так и пищеварения за счет переваривания фагоцитированных пищевых частиц, захваченных из воды.
Осуществление одной и той же клеткой функций движения и пищеварения малоэффективно. С этим связана последующая специализация клеток в направлении преимущественно пищеварения или обеспечения движения. Результатом является возникновение фагоцитобласта (внутреннего слоя амебовидных клеток, занимающихся пищеварением) и кинобласта (наружного слоя клеток со жгутиками, обеспечивающими движение).
Стойкая дифференцировка соматических клеток по функциям и строению, возникшая первоначально на фоне выделения двух клеточных слоев, явилась ключевым моментом в происхождении многоклеточных. Именно с двуслойностью связано появление жидкой внутренней среды, через которую клетки обмениваются химическими сигналами, а также дальнейшее обособление и специализация части поверхностных клеток в направлении восприятия внешних раздражителей и передача возбуждения на другие клетки, располагающиеся в отдалении от них. Таким образом возникают предпосылки к формированию нервной системы.
Гипотетический предок многоклеточных животных назван фагоцителлой (рис. 13.11). Он плавал в толще воды за счет биения ресничек кинобласта, а питался, захватывая взвешенные в среде частички пищи и переваривая их клетками фагоцитобласта. На более поздних этапах эволюции происходили многочисленные адаптации потомков фагоцителлы к многообразным условиям существования при оседании их на дно или при перемещении к поверхности, а также при изменении источников питания (захват мелких или крупных, живых или мертвых пищевых частиц).
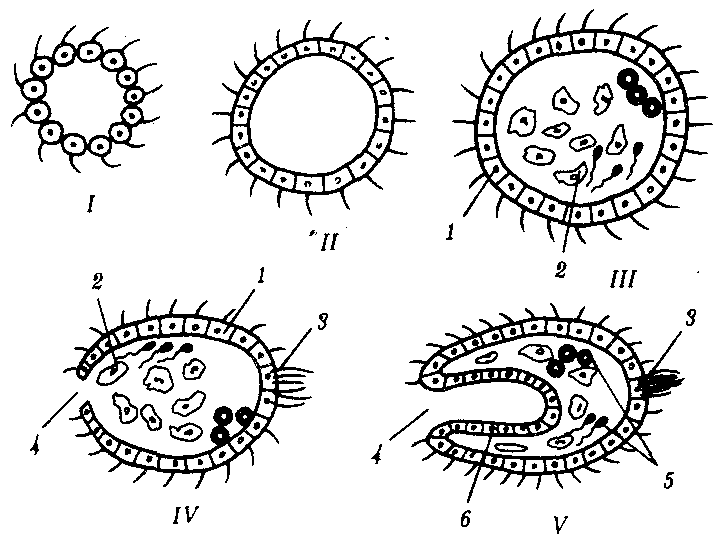
Рис. 13.11. Этапы происхождения многоклеточности: I, II—сферические колонии жгутиковых, III—V—фагоцителлы разной степени сложности; 1—кинобласт, 2—рыхлый фагоцитобласт, 3—скопление чувствительных клеток на переднем конце тела, 4—ротовое отверстие, 5—половые клетки, 6—эпителизованный фагоцитобласт
Большое значение в эволюции потомков фагоцителлы имели также изменения характера движения: пассивное движение или прикрепленный образ жизни обусловливают лучевой тип симметрии, в то время как активное перемещение в определенном направлении предусматривает формирование двубоковой, или билатеральной, симметрии. В результате возникло огромное многообразие форм многоклеточных животных.
Представления о происхождении многоклеточных, изложенные здесь, являются развитием гипотезы И.И. Мечникова (1877—1880) о том, что многоклеточность возникла на базе шаровидных колоний жгутиковых, часть клеток которых иммигрировала внутрь для переваривания пищевых частиц, в результате чего оформилась дифференцировка на экто- и энтодерму.
13.5.3. Основные этапы прогрессивной эволюции многоклеточных животных
Важным шагом в эволюции многоклеточных животных явилось возникновение третьего зародышевого листка — мезодермы. Мезодерма обеспечивает возможность дифференцировки мышечной, соединительной тканей и скелета, а также многоклеточных половых желез, в которых созревающие гаметы оказываются надежно защищены от неблагоприятных средовых воздействий. Практически все трехслойные животные ведут активно подвижный образ жизни, благодаря чему приобретают билатеральный тип симметрии. Вместе с тем у трехслойных животных с интенсивным обменом веществ, активно перемещающихся с помощью мышц, возникают проблемы с выведением большого количества продуктов диссимиляции из тканей — производных мезодермы, в то время как эктодермальные и энтодермальные клетки выделяют их за счет диффузии соответственно либо наружу, либо в просвет пищеварительной полости. Поэтому именно у трехслойных впервые появляется и прогрессивно эволюционирует

выделительная система.
Следующий значительный этап эволюции животных — возникновение вторичной полости тела, или целома, первоначально функционирующего как гидростатический скелет, а также выполняющего половую и выделительную функции в связи с тем, что продукты диссимиляции и половые клетки попадают в целом и только потом выделяются наружу (см. § 14.5).
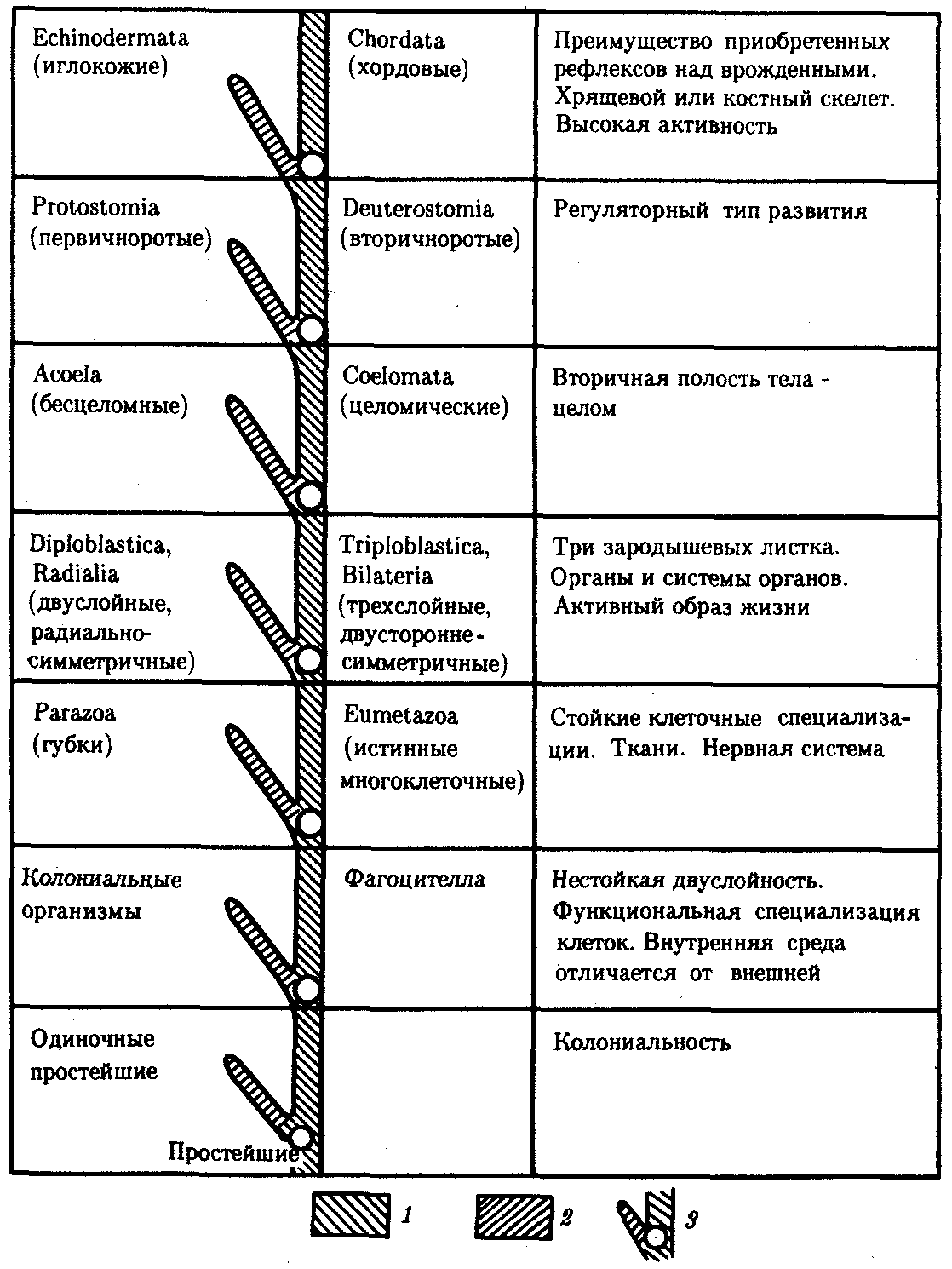
Рис. 13.12. Главные направления эволюции групп в животном царстве:
1—прогрессивное направление, 2—адаптивное направление, 3—узловые моменты в прогрессивной эволюции; каждому узловому моменту соответствует его характеристика, обозначенная в правом столбце
Существенным этапом дальнейшей эволюции многоклеточных является возникновение регуляторного типа эмбрионального развития (см. разд. 8.3.1 и 8.3.2), в результате которого в развивающемся зародыше доминирует целостность морфогенетических процессов над их составляющими. Благодаря этому зародыш развивается относительно автономно в соответствии со своей генетической программой и способен компенсировать даже серьезные повреждения. Организмы, характеризующиеся такими особенностями, относят к группе вторичноротых, в отличие от первичноротых, у которых эмбриональное развитие протекает по мозаичному типу (см. разд. 8.3.1).
Наиболее крупные систематические группировки в царстве Животные называют типами. За период существования жизни на Земле их было не менее 35. К настоящему времени некоторые из них вымерли; сейчас на Земле обитают животные 26 типов.
На рис. 13.12 приведена схема главных направлений эволюции в Животном царстве, а на рис. 13.13 изображены представители основных типов современных животных. Интересно, что к концу протерозойской эры (2,7 млрд. лет тому назад) на Земле уже существовали представители всех типов животного мира и основные узловые моменты в прогрессивной эволюции животных ими были пройдены.
Существование разнообразных живых организмов на Земле на протяжении около 3 млрд. лет, а также возникновение человека как биосоциального существа определяет в настоящее время картину современного органического мира.
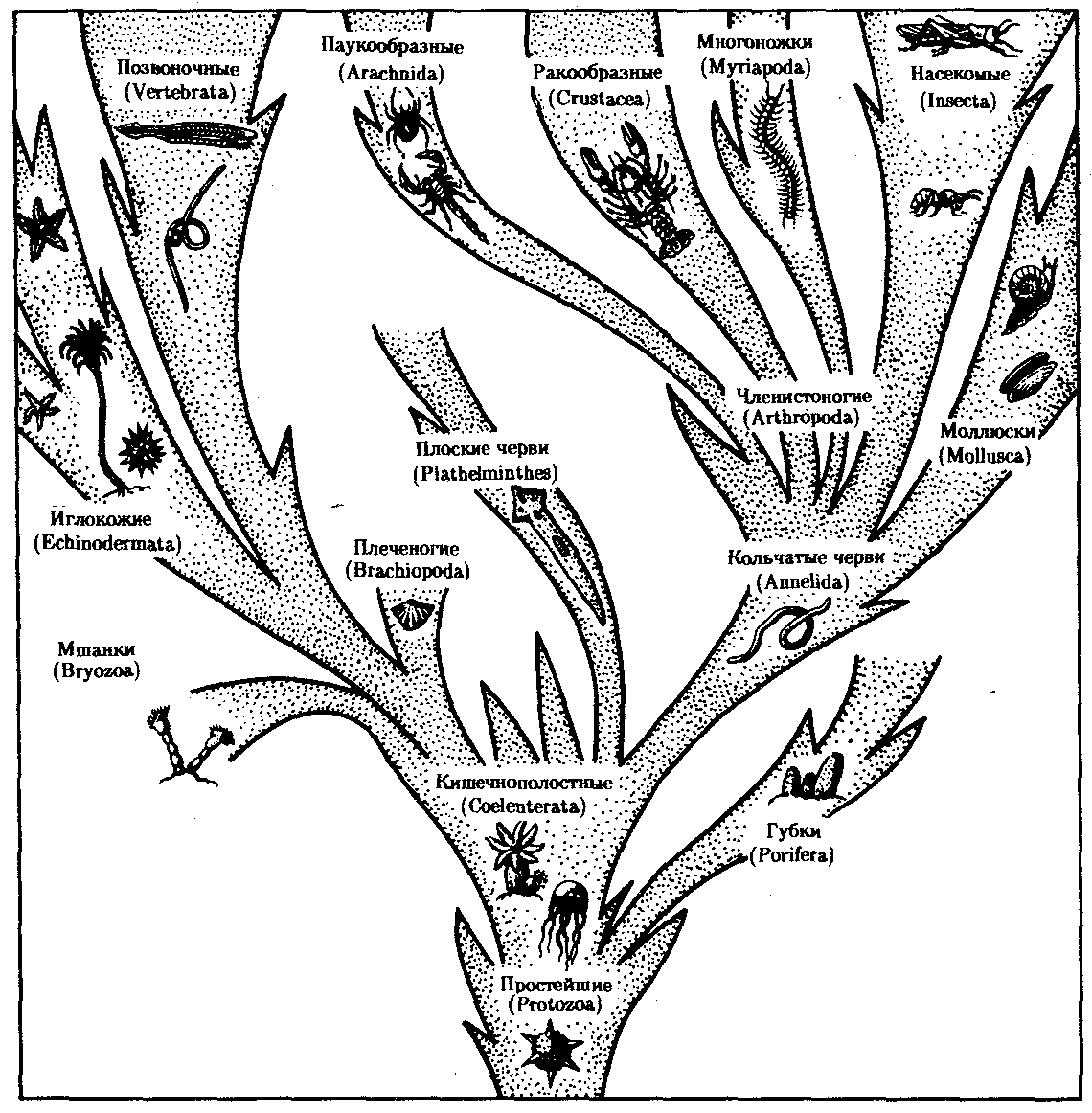
Рис. 13.13. Основные типы животного царства и их филогенетические взаимоотношения
13.5.4. Характеристика типа Хордовые
Хордовые представляют собой наиболее высокоорганизованный тип в животном мире. Их общими чертами являются следующие.
1. Внутренний осевой скелет представлен хордой, которая присутствует в эмбриогенезе у всех представителей типа, а у высших дополняется, а затем и замещается позвоночником.
2. Над хордой располагается центральная нервная система в виде нервной трубки с полостью — невроцелем.
3. В боковых стенках глотки находятся жаберные щели, соединяющие ее полость с внешней средой. У рыб и некоторых земноводных они сохраняются в течение всей жизни, у высших хордовых — только в эмбриональном периоде.
4. Тело построено метамерно. У низших хордовых и у зародышей высших сегментация распростраянется на все системы органов, у высших ярко выражена только в эмбриональном периоде. Позже частично сохраняется только в опорно-двигательном аппарате, нервной и кровеносной системах.
5. Органами поддержания равновесия и движения являются конечности, причем у низших хордовых большее значение имеют непарные, а у высших — парные.
6. Общий план строения хордовых представлен на рис. 13.14. На спинной стороне расположена нервная трубка, под ней — хорда или заменяющий ее позвоночник. Глубже находится пищеварительная трубка с развивающейся из нее дыхательной системой, а под ней — вентральный пульсирующий кровеносный сосуд или сердце. По бокам от нервной трубки и хорды лежат сомиты, а по бокам от кишки — спланхнотомы, внутри которых расположен целом.
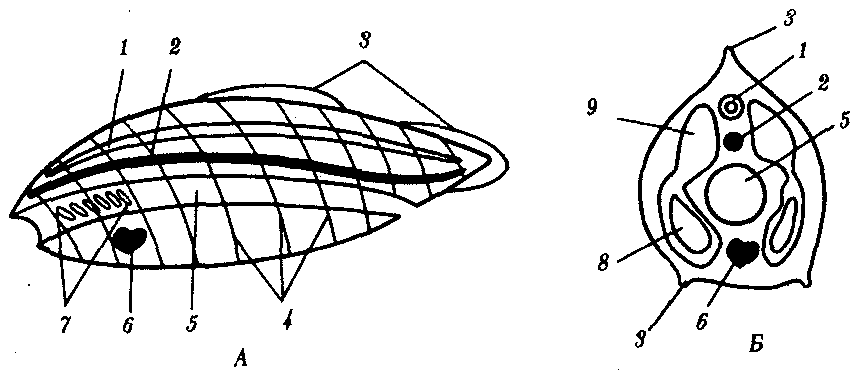
Рис. 13.14. Общий план строения хордового животного. А — вид сбоку; Б — поперечный срез:
1—нервная трубка, 2—хорда, 3—плавники, 4—сегменты тела, 5—пищеварительная трубка, 6—пульсирующий сосуд или сердце. 7—жаберные щели в глотке, 8—целом, 9—сомит
13.5.5. Систематика типа Хордовые
Из четырех подтипов хордовых — Полухордовые Hemichordata, Личиночно-хордовые Urochordata, Бесчерепные Acrania и Позвоночные Vertebrata — остановимся на двух последних, имеющих отношение к прогрессивному направлению в эволюции этого типа животных.
Подтип Бесчерепные состоит лишь из одного класса — Головохордовые Cephalochordata, к которому относится ланцетник; подтип Позвоночные включает следующие классы: Круглоротые Cyclostomata, Хрящевые рыбы Chondrichthyes, Костные рыбы Osteichthyes, Земноводные Amphibia, Пресмыкающиеся Reptilia, Птицы Aves и Млекопитающие Mammalia.
13.5.6. Подтип Бесчерепные Acrania
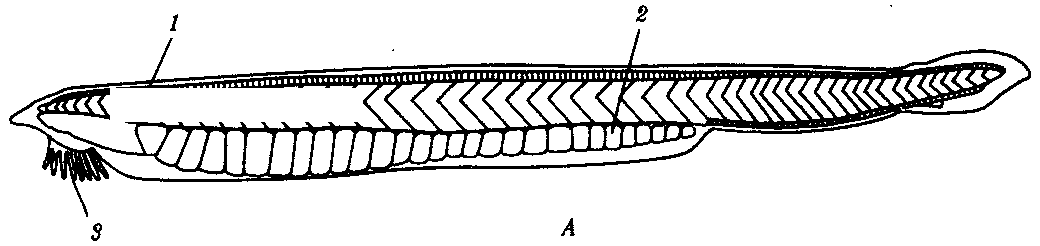
Характерной особенностью животных этого подтипа, представителем которого является ланцетник Branchiostoma lanceatum (рис. 13.15), является малоподвижный придонный образ жизни и пассивное питание за счет фильтрации воды. Тело ланцетника полупрозрачно, покрыто однослойным цилиндрическим эпителием. Дерма развита слабо. Хорда тянется от головного до хвостового конца, а нервная трубка замкнута на спинной стороне неполно. В ней расположены светочувствительные глазки Гессе, обеспечивающие ланцетнику лишь восприятие света и темноты.
Около половины длины пищеварительной трубки составляет глотка, которая начинается позади ротовой полости и пронизана более чем 100 парами жаберных щелей, ведущих в околожаберную полость. Последняя открывается во внешнюю среду на брюшной стороне тела. Кишка в передней ее части снабжена печеночным выростом, гомологичным печени других хордовых, и заканчивается анальным отверстием. При поступлении воды через рот в глотку и далее в околожаберную полость осуществляется как отцеживание взвешенных в воде пищевых частиц, так и газообмен в кровеносных сосудах межжаберных перегородок.
Кровеносная система замкнута. Имеется один круг кровообращения, функцию сердца выполняет пульсирующая брюшная аорта.
Органы выделения — метамерно расположенные на межжаберных перегородках нефридии, состоящие из воронок, которые собирают продукты диссимиляции из целома и выводят их в околожаберную полость.
Метамерно организованы и половые железы, выделяющие гаметы по мере их созревания также в околожаберную полость и далее во внешнюю среду вне зависимости от наличия поблизости половозрелых организмов противоположного пола.
Данные сравнительной анатомии и эмбриологии позволяют сделать вывод о том, что предками бесчерепных были мелкие свободнопла-вающие двусторонне-симметричные существа с сегментированной мускулатурой и с небольшим количеством жаберных щелей, открывающихся наружу. Так же как и у ланцетника, питание их было пассивным и осуществлялось за счет фильтрации воды через жаберные щели, а половые продукты периодически выбрасывались в воду. Эти первичные бесчерепные дали начало двум ветвям в эволюции хордовых. Представители одной из них перешли к придонному образу жизни. От них произошли предки современных бесчерепных. Другая ветвь характеризовалась усилением двигательной активности. Арогенная эволюция в этой ветви хордовых привела к формированию подтипа Позвоночные.