ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 03.02.2024
Просмотров: 1099
Скачиваний: 0

СОДЕРЖАНИЕ
ПОПУЛЯЦИОННО-ВИДОВОЙ УРОВЕНЬ ОРГАНИЗАЦИИ ЖИЗНИ
ВИДООБРАЗОВАНИЕ В ПРИРОДЕ. ЭЛЕМЕНТАРНЫЕ ЭВОЛЮЦИОННЫЕ ФАКТОРЫ
11.5. ГЕНЕТИКО-АВТОМАТИЧЕСКИЕ ПРОЦЕССЫ (ДРЕЙФ ГЕНОВ)
11.7. НАСЛЕДСТВЕННЫЙ ПОЛИМОРФИЗМ ПРИРОДНЫХ ПОПУЛЯЦИЙ.
11.9. ПРОИСХОЖДЕНИЕ БИОЛОГИЧЕСКОЙ ЦЕЛЕСООБРАЗНОСТИ
ДЕЙСТВИЕ ЭЛЕМЕНТАРНЫХ ЭВОЛЮЦИОННЫХ ФАКТОРОВ
12.1. ПОПУЛЯЦИЯ ЛЮДЕЙ. ДЕМ, ИЗОЛЯТ
12.2. ВЛИЯНИЕ ЭЛЕМЕНТАРНЫХ ЭВОЛЮЦИОННЫХ ФАКТОРОВ НА ГЕНОФОНДЫ ЧЕЛОВЕЧЕСКИХ ПОПУЛЯЦИЙ
12.3. ГЕНЕТИЧЕСКОЕ РАЗНООБРАЗИЕ
13.1. ЭВОЛЮЦИЯ ГРУПП ОРГАНИЗМОВ
13.2. СООТНОШЕНИЕ ОНТО- И ФИЛОГЕНЕЗА
13.4. Организм как целое в историческом
Соотносительные преобразования органов
13.5. СОВРЕМЕННАЯ СИСТЕМА ОРГАНИЧЕСКОГО МИРА
ФИЛОГЕНЕЗ СИСТЕМ ОРГАНОВ ХОРДОВЫХ
14.2. ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
И ДАЛЬНЕЙШАЯ ЭВОЛЮЦИЯ ЧЕЛОВЕКА
15.3. ХАРАКТЕРИСТИКА ОСНОВНЫХ ЭТАПОВ АНТРОПОГЕНЕЗА
15.4. ВНУТРИВИДОВАЯ ДИФФЕРЕНЦИАЦИЯ ЧЕЛОВЕЧЕСТВА
БИОГЕОЦЕНОТИЧЕСКИЙ УРОВЕНЬ ОРГАНИЗАЦИИ ЖИЗНИ
16.1. БИОГЕОЦЕНОЗ - ЭЛЕМЕНТАРНАЯ ЕДИНИЦА БИОГЕОЦЕНОТИЧЕСКОГО УРОВНЯ ОРГАНИЗАЦИИ ЖИЗНИ
17.2. ЧЕЛОВЕК КАК ОБЪЕКТ ДЕЙСТВИЯ ЭКОЛОГИЧЕСКИХ ФАКТОРОВ.
АДАПТАЦИЯ ЧЕЛОВЕКА К СРЕДЕ ОБИТАНИЯ
17.4. РОЛЬ АНТРОПОГЕННЫХ ФАКТОРОВ
В ЭВОЛЮЦИИ ВИДОВ И БИОГЕОЦЕНОЗОВ
МЕДИЦИНСКАЯ ПАРАЗИТОЛОГИЯ. ОБЩИЕ ВОПРОСЫ
18.2. ФОРМЫ МЕЖВИДОВЫХ БИОТИЧЕСКИХ СВЯЗЕЙ В БИОЦЕНОЗАХ
18.3. КЛАССИФИКАЦИЯ ПАРАЗИТИЗМА
18.4. РАСПРОСТРАНЕННОСТЬ ПАРАЗИТИЗМА
18.5. ПРОИСХОЖДЕНИЕ ПАРАЗИТИЗМА
18.6. АДАПТАЦИИ К ПАРАЗИТИЧЕСКОМУ ОБРАЗУ ЖИЗНИ. ОСНОВНЫЕ ТЕНДЕНЦИИ
18.8. ФАКТОРЫ ВОСПРИИМЧИВОСТИ ХОЗЯИНА К ПАРАЗИТУ
18.9. ДЕЙСТВИЕ ХОЗЯИНА НА ПАРАЗИТА
18.10. СОПРОТИВЛЕНИЕ ПАРАЗИТОВ РЕАКЦИЯМ ИММУНИТЕТА ХОЗЯИНА
18.11. ВЗАИМООТНОШЕНИЯ В СИСТЕМЕ ПАРАЗИТ - ХОЗЯИН НА УРОВНЕ ПОПУЛЯЦИЙ
18.12. СПЕЦИФИЧНОСТЬ ПАРАЗИТОВ ПО ОТНОШЕНИЮ К ХОЗЯИНУ
18.13. ПРИРОДНО-ОЧАГОВЫЕ ЗАБОЛЕВАНИЯ
19.2. Простейшие, обитающие в полостных органах, сообщающихся с внешней средой
19.3. Простейшие, обитающие в тканях
19.4. ПРОСТЕЙШИЕ — ФАКУЛЬТАТИВНЫЕ ПАРАЗИТЫ ЧЕЛОВЕКА
20.1. ТИП ПЛОСКИЕ ЧЕРВИ PLATHELMINTHES
20.2. ТИП КРУГЛЫЕ ЧЕРВИ NEMATHELMINTHES
21.1. КЛАСС ПАУКООБРАЗНЫЕ ARACHNOIDEA
ЭВОЛЮЦИЯ ПАРАЗИТОВ И ПАРАЗИТИЗМА ПОД ДЕЙСТВИЕМ АНТРОПОГЕННЫХ ФАКТОРОВ
23.1. ПРОИСХОЖДЕНИЕ ЯДОВИТОСТИ
23.2. ЧЕЛОВЕК И ЯДОВИТЫЕ ЖИВОТНЫЕ
24.1. СОВРЕМЕННЫЕ КОНЦЕПЦИИ БИОСФЕРЫ
24.2. СТРУКТУРА И ФУНКЦИИ БИОСФЕРЫ
25.2. ПУТИ ВОЗДЕЙСТВИЯ ЧЕЛОВЕЧЕСТВА


Рис. 13.15. Строение ланцетника. А—общий вид; Б—сагиттальный срез; В—поперечный срез:
1—миомер, 2—гонада, 3—щупальца предротовой воронки, 4—спинной мозг, 5—хорда, 6—жаберные щели, 7—лучи (камеры) спинного плавника, 8— анальное отверстие, 9—кишечник, 10—атриопор, 11—печеночный вырост, 12—корень спинной аорты, 13—наджаберная бороздка, 14—глотка, 15—жаберная перегородка, 16—атриальная полость, 17—металлевральная складка, 18—эндостиль
13.5.7. Подтип Позвоночные Vertebrata
Основные особенности образа жизни позвоночных — их активные перемещения в пространстве, обеспечивающие им эффективное разыскивание пищи и представителей противоположного пола при размножении. Это достигается следующими ароморфными чертами их организации: усовершенствованный опорно-двигательный аппарат (сегментированный позвоночник взамен хорды); конечности с собственными мышцами и скелетом; дифференцировка головы с черепом, содержащим головной мозг (состоит из пяти отделов); развитые органы чувств. Интенсификации обмена веществ способствует выделение специализированных органов дыхания — жабр или легких, а также дифференцировка сердца из брюшного кровеносного сосуда. Выведение продуктов диссимиляции осуществляется специализированным компактным органом — почкой. Половые продукты выделяются гонадами позвоночных только в период размножения в момент встречи особей противоположных полов (рис. 13.16).

Рис. 13.16. Строение позвоночного челюстноротого животного:
1—череп, 2—головной мозг, 3—позвоночник, 4—спинной мозг, 5—головная почка, 6—туловищная почка, 7—половая железа с протоком, 8—тазовая почка, 9—клоака, 10—мочевой пузырь, 11—задняя парная конечность, 12— кишка, 13 — селезенка, 14—поджелудочная железа, 15—печень, 16—желудок, 17— передняя парная конечность, 18—легкое, 19—сердце, 20—глотка, 21—жаберные дуги, 22—жаберные щели, 23—челюсти
Семь перечисленных выше классов типа позвоночных фактически являются ступенями, соответствующими поэтапному повышению уровня организации в этом филогенетическом стволе эволюционного древа животного мира (рис. 13.17).
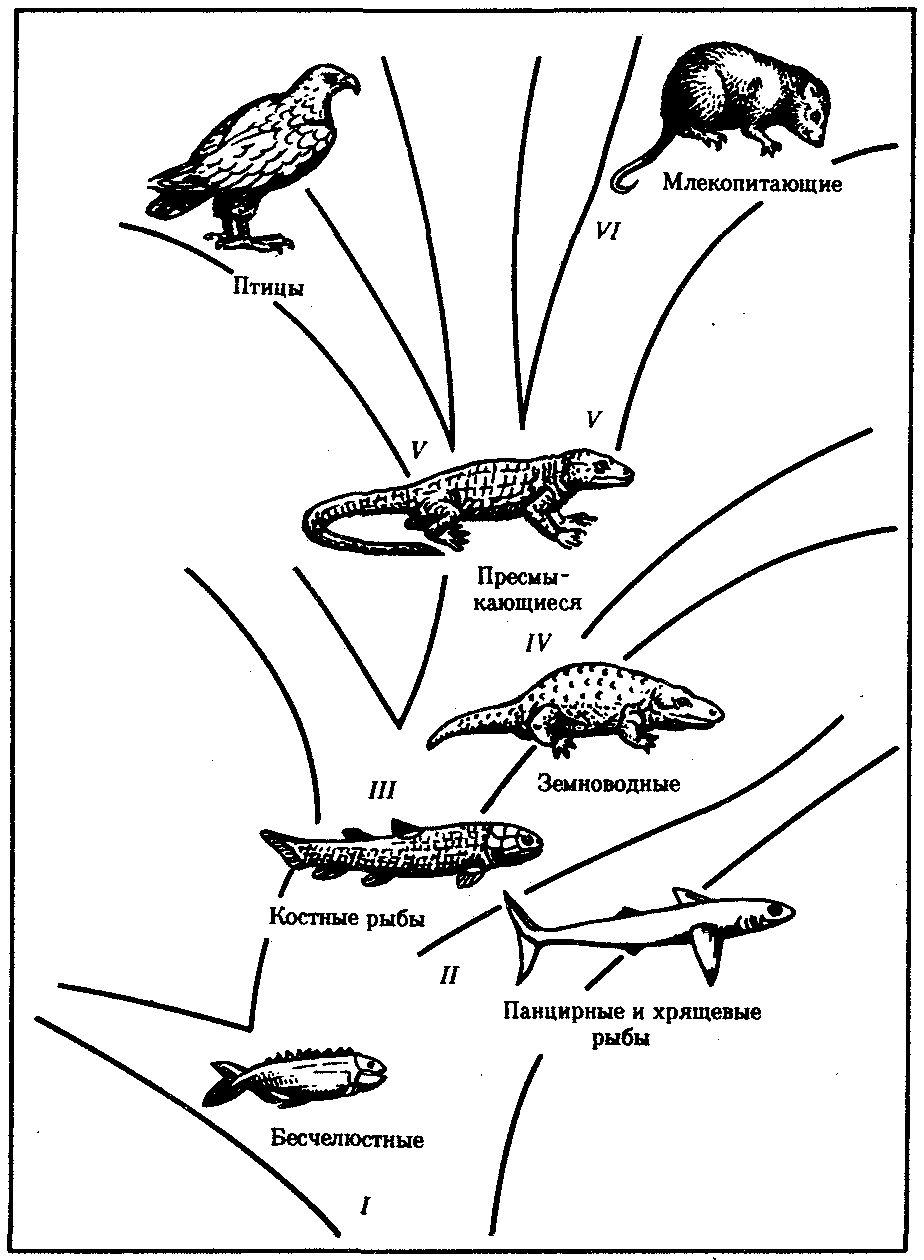
Рис. 13.17. Узловые моменты в прогрессивной эволюции хордовых:
I—появление хрящевого скелета, дифференцировка центральной нервной системы на головной и спинной мозг, II—появление челюстей, III—появление парных конечностей наземного типа и легких, IV—преодоление барьера влажности, V—теплокровность, VI—внутриутробное развитие
Класс Круглоротые Cyclostomata — наиболее архаичный среди позвоночных. Представители его смогли дожить до настоящего времени в основном благодаря переходу к паразитическому образу жизни. Представителями являются миноги. Главная их особенность — неподвижность рта, которая обусловлена особым строением жаберных дуг, выполняющих только функцию опоры для глотки. Имеются непарные спинные и хвостовые плавники.
Активизация питания — активный захват пищи — важнейшее ароморфное приобретение надкласса Рыбы Pisces. Благодаря этому в отличие от бесчелюстных круглоротых рыбы и все более высокоорганизованные позвоночные относятся к группе Челюстноротые Gnathostomata. Челюсти возникли в результате преобразования передних жаберных дуг (см. разд. 14.2.1). Кроме того, у рыб прогрессивно развиваются парные конечности с собственным скелетом, а у наиболее высокоорганизованных из них — и плавательный пузырь, выполняющий гидростатические функции.
Следующий этап в прогрессивной эволюции позвоночных — выход на сушу, обеспечивающийся дифференцировкой четырех парных конечностей и утратой значения хвостового плавника как органа движения у земноводных. В связи с этим земноводные вместе с более прогрессивными позвоночными объединяются в группу Четвероногие Tetrapoda. Выход на сушу сопровождается преобразованием плавательного пузыря в легкие и появлением в связи с этим двух кругов кровообращения и трехкамерного сердца.
Пресмыкающиеся Reptilia успешно преодолели барьер влажности. Они имеют сухую кожу, предохраняющую организм от высыхания, и стали независимыми от водной среды даже при размножении, приобретя способность к внутреннему оплодотворению и откладке яиц с плотными оболочками и запасами жидкости. Амниотическая оболочка их зародышей, формирующаяся в ходе эмбриогенеза, создает благоприятные условия для их развития вне зависимости от влажности окружающей среды. Поэтому пресмыкающиеся вместе с птицами и млекопитающими относятся к группе Амниоты Amniota, или Первичноназемные животные.

Ключевой адаптацией птиц и млекопитающих является их теплокровность, или гомойотермность, обеспечивающая резкий биологический прогресс этих классов благодаря независимости от климатических факторов среды. Эти классы объединяются в группу Гомойотермные.
Наконец, Плацентарные млекопитающие Placentalia характеризуются внутриутробным эмбриональным развитием, в еще меньшей степени, чем у пресмыкающихся, зависящим от окружающей среды. Выкармливание потомства молоком значительно повышает шансы на выживание.
Кроме того, преимущественное развитие в центральной нервной системе коры больших полушарий переднего мозга обеспечивает кардинальное усложнение поведения на фоне превалирования условных рефлексов над безусловными и формирования сложных инстинктов. Биологическая организация наиболее прогрессивного отряда млекопитающих — Приматы Primates — явилась предпосылкой происхождения человека именно в рамках этого отряда.
ГЛАВА 14
ФИЛОГЕНЕЗ СИСТЕМ ОРГАНОВ ХОРДОВЫХ
Филогенез систем органов хордовых рассмотрен в соответствии с прогрессивным направлением эволюции этого типа животных от подтипа Бесчерепные до класса Млекопитающие. Организация систем органов класса Птицы не описана в связи с тем, что птицы произошли от пресмыкающихся значительно позже млекопитающих и являются боковой ветвью эволюции хордовых.
14.1. Наружные покровы
Покровы любых животных всегда выполняют функцию восприятия внешних раздражении, а также защищают тело от вредных воздействий среды. Интенсификация первой функции покровов приводит в процессе эволюции многоклеточных животных к возникновению нервной системы и органов чувств. Интенсификация второй функции сопровождается дифференцировкой. Характерно также расширение функций, в результате чего кожа как орган защиты участвует также в газообмене, терморегуляции и выделении, выкармливании потомства. Это связано с усложнением строения слоев кожи, появлением и дальнейшим преобразованием многочисленных придатков и желез.
У всех хордовых кожа имеет двойное — экто- и мезодермальное — происхождение. Из эктодермы развивается эпидермис, из мезодермы — дерма. Для бесчерепных характерна слабая степень дифференцированности обоих слоев кожи. Эпидермис однослойный цилиндрический, содержащий одноклеточные слизистые железы, дерма рыхлая, содержит небольшое количество соединительнотканных клеток.
В подтипе Позвоночные эпидермис становится многослойным, причем в нижнем слое клетки постоянно размножаются, а в верхних слоях — дифференцируются, гибнут и слущиваются. В дерме появляются соединительнотканные волокна, придающие покровам прочность. Кожа образует придатки, многообразные в зависимости от образа жизни и уровня организации, а также железы, выполняющие различные функции.
У рыб в эпидермисе железы одноклеточные. Как и у ланцетника, они выделяют слизь, облегчающую движения в воде. Тело рыб покрыто чешуей, имеющей разное строение в зависимости от их систематического положения. Чешую хрящевых рыб называют плакоидной. Она имеет форму шипа и состоит из дентина, покрытого снаружи эмалью (рис. 14.1). Дентин мезодермального происхождения
, он образуется за счет функционирования соединительнотканных клеток, выпячивающихся снаружи в виде сосочка. Эмаль, представляющая собой более твердое, чем дентин, неклеточное вещество, формируется сосочком эпидермиса и покрывает плакоидную чешую снаружи.
Вся поверхность тела хрящевых рыб, а также ротовая полость, слизистая оболочка которой происходит из эктодермы, покрыты плакоидной чешуей. Естественно, что функции чешуи в ротовой полости связаны с захватом и удержанием пищи, поэтому они сильно увеличены и являются зубами. У костных рыб чешуя иного типа. Она имеет вид тонких круглых костных пластинок, покрытых тонким слоем эпидермиса. Костная чешуя развивается полностью за счет дермы, но по происхождению связана с примитивной плакоидной.
Кожа примитивных вымерших земноводных — стегоцефалов — соответствовала покровам рыб и тоже была покрыта чешуей. Современные амфибии имеют тонкую гладкую кожу без чешуи, принимающую участие в газообмене. Этому способствует наличие большого количества многоклеточных слизистых желез, секрет которых постоянно увлажняет покровы и обладает бактерицидными свойствами. Некоторые кожные железы ряда земноводных дифференцировались в органы-продуценты токсинов, защищающих их от врагов (см. разд. 23.1).
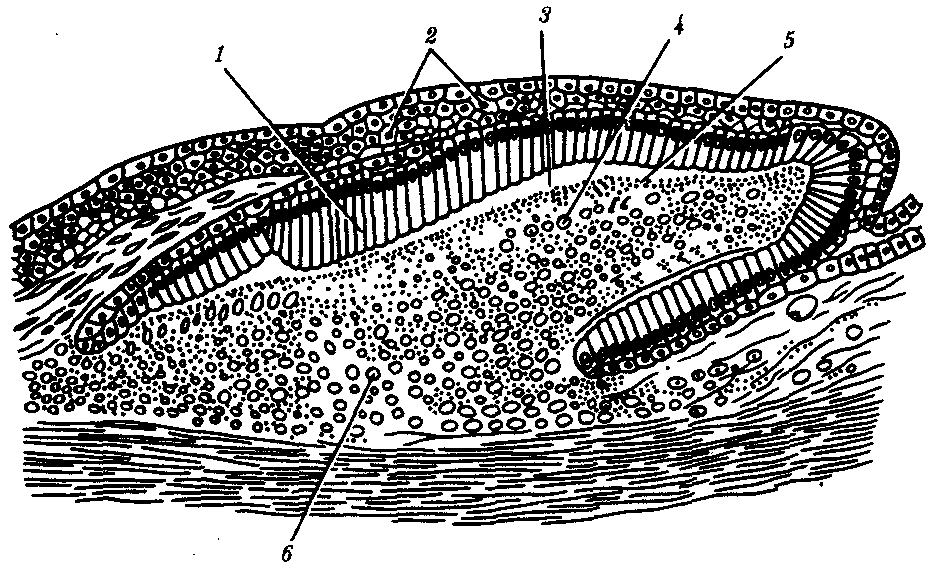
Рис. 14.1. Закладка плакоидной чешуи:
1—клетки-эмалеобразователи, 2—эпидермис, 3—эмаль, 4—склеробласты—дентинообразователи, 5—дентин, 6—сосочек дермы
Пресмыкающиеся, перешедшие полностью к наземному существованию, имеют сухую кожу, не участвующую в дыхании. Верхний слой эпидермиса ороговевает. Роговые чешуйки у некоторых рептилий тонки и эластичны, у других—сливаются вместе, образуя, как у черепах, мощный роговой панцирь. Большинство пресмыкающихся по мере роста линяют, сбрасывая периодически свой роговой покров. Современные пресмыкающиеся не имеют кожных желез.
Кожные покровы млекопитающих построены наиболее сложно в связи с выполнением ими многообразных функций. Характерны различные производные кожи: волосы, когти, рога, копыта, а также потовые, сальные и млечные железы. Более примитивные млекопитающие — насекомоядные, грызуны и некоторые другие — наряду с волосяным покровом сохранили также роговые чешуи на хвосте. Волосы у них растут в промежутках между чешуйками, группами по 3—7. У более прогрессивных млекопитающих, утративших чешуи, сохраняется такое же расположение волос (рис. 14.2),