ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 19.10.2024
Просмотров: 25
Скачиваний: 0
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
СОДЕРЖАНИЕ
12. Химический состав и строение клеточной стенки, ее функциональное значение
34. Роль активной деятельности цитоплазмы и поглощении воды клеткой
54. Нарушение водообмена, его причины и последствия
89. Связь дыхания и брожения. Пути окисления пировиноградной кислоты в растительных тканях
95. Дегидрогеназы, их химическая природа и характер действия
130. Ингибиторы их физиологическая роль и применение в практике
176. Физиологические особенности засухоустойчивых сельскохозяйственных условий
Большая часть присутствующей в клетке воды находится в вакуоле. Там же содержится большое количество других веществ— сахаров, органических кислот, солей. Для того чтобы попасть в вакуоль, вода должна пройти сквозь стенку клетки, а также через две мембраны и лежащую между ними цитоплазму. Обычно эти три структуры рассматривают совместно как единый мембранный барьер, обладающий свойствами полупроницаемости, т. е. проницаемостью только для воды. Согласно молекулярно-кинетической теории, молекулы всех веществ находятся в состоянии быстрого хаотического движения, скорость которого зависит от энергии этих молекул. Молекулы воды малы и проходят через клеточные мембраны намного быстрее, чем молекулы других веществ. Они диффундируют во всех направлениях: в клетку и из клетки, в различные клеточные органеллы и из них. Движение воды осуществляется по градиенту ее активности„ которая характеризуется водным потенциалом. Молекулы содержащихся в вакуоле растворенных веществ снижают активность молекул воды в клетке. Вследствие этого кинетическая энергия воды в вакуоле ниже, чем кинетическая энергия относительно более чистой воды вне клетки. В результате внутрь клетки в вакуоль проникает больше молекул воды, чем выходит из нее. Объем вакуоли увеличивается и создается тургор, в результате которого содержимое клетки прижимается к ее стенке.
Диффузия воды через полупроницаемую мембрану называется осмосом. Если два раствора разделены полупроницаемой мембраной, то вода из раствора с меньшей концентрацией веществ (ги-потонического) будет диффундировать в раствор с большей концентрацией веществ (гипертонический). Диффузия будет происходить до тех пор, пока концентрации веществ в обоих растворах не станут одинаковыми (изотоническими).
Если в осмометре раствор сахарозы отделить от чистой воды полупроницаемой мембраной (рис. 8.2), то молекулы воды в силу диффузии будут перемещаться из чистой воды в раствор сахарозы, где концентрация воды меньше, т.е. от высокого водного потенциала к более низкому. Это приводит к увеличению объема раствора сахарозы и подъему жидкости в трубке осмометра. При этом повышается гидростатическое давление, под которым находится раствор в осмометре. При некоторой высоте столба раствора в трубке скорости диффузии воды сравняются и подъем его прекратится.
Наибольшее давление, которое способен преодолеть данный раствор, всасывая воду через полупроницаемую мембрану, является его осмотическим давлением. Осмотический потенциал раствора прямо зависит от концентрации растворенного вещества и по величине равен потенциалу гидростатического давления, по противоположен ему по знаку. Он всегда имеет отрицательное значение. Величина осмотического потенциала неоднозначна у разных видов растений и в разных частях одного и того же растения. Для вакуолярного сока клеток корней она составляет 12 -13 ат., а для клеток надземных органов 10 – 26 ат. Это обуславливает вертикальный градиент асмотического потенциала и сосущей силы от корней к листьям. У растений- галофитов, растущих в условиях засоления, осмотический потенциал очень высок, достигая 150 ат.
Силу, с которой вода входит в клетку, называют сосущей силой (5), она тождественна водному потенциалу клетки. Величина сосущей силы определяется осмотическим давлением клеточного сока (z) и тургорным (гидростатическим) давлением в клетке
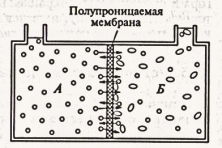
Рис. 8.2. Схема простейшего осмометра:
1 — сосуд с водой; Б — сосуд с 10%-м
раствором сахарозы (кружки — молекулы воды, овалы — молекулы сахарозы).
(Р), которое равно противодавлению клеточной стенки, возникающему при ее эластичном растяжении: S = к — Р.
В условиях разной оводненности соотношение между всеми компонентами этого уравнения меняется. Когда клетка полностью насыщена водой (полностью тургесцентна), ее сосущая сила равна нулю, а тургорное давление равно потенциальному осмотическому: S= 0; Р= к. Состояние полного тургора наблюдается в клетках водных растений. Клетки наземных растений из-за транспирации почти никогда не бывают полностью насыщенными во-дой. Если подача воды к клетке уменьшается (при усилении вет-ра, при недостатке влаги в почве и т.д.), то вначале возникает водный дефицит в клеточных стенках, водный потенциал которых становится ниже, чем в вакуолях, и вода начинает перемещаться в клеточные стенки. Отток воды из вакуоли снижает тур-горное давление в клетках и, следовательно, увеличивает их сосущую силу. При длительном недостатке влаги большинство клеток теряет тургор и растение завядает. В этих условиях

Р = 0; S = к.
Обезвоживание клетки называется плазмолизом.
Явление плазмолиза можно наблюдать, помещая кусочки ткани в гипертонический раствор. Отток воды из клеток приводит к уменьшению объема протопластов и их отделению от клеточных стенок. Пространство между клеточной стенкой и сократившимся протопластом заполняет наружный раствор, который легко проходит через клеточную стенку, практически не препятствующую движению воды. Клетку в таком состоянии называют плазмолизированной. При помещении клетки в чистую воду она может восстановить свой тургор. Чрезмерный дефицит приводит к разрыву протопластов и гибели клеток.
В условиях водного дефицита в молодых тканях резкое усиление потери воды, например, при суховее, приводит к потере тургора и протопласты, сокращаясь в объеме, не отделяются от клеточных стенок, а тянут их вслед за собой. Клетки и ткани сжимаются. Это явление называется циторризом.
54. Нарушение водообмена, его причины и последствия
Нарушение водообмена у растений происходит при недостаточном водоснабжении, то есть, при наступлении засухи.
Засуха - неблагоприятное сочетание метеорологических условий, которые не обеспечивают потребностей растения в воде. Засуха бывает почвенная и атмосферная. Причины почвенной засухи - отсутствие дождя на протяжении длительного времени, испарение с поверхности почвы и транспирация, что приводит к высушиванию корнеобитаемого слоя почвы. Последнюю высушивают также сильные ветры. При почвенной засухе влажность воздуха бывает пониженной. Атмосферная засуха вызывается массами сухого и нагретого воздуха (суховея). Засуха, как почвенная, так и атмосферная, задерживает рост растений, уменьшает их листовую поверхность, в результате чего урожай снижается.
Влияние почвенной (обезвоживание) и атмосферной (перегрев) засух проявляется многими признаками. Под влиянием обезвоживания нарушается синтетическая способность растений, наблюдается распад белков и изменяется коллоидно-химическое состояние цитоплазмы, снижается количество накопленного растением органического вещества из-за задержки роста листьев и уменьшается их рабочая поверхность. Особенно чувствительны к засухе листья, находящиеся в фазе эмбрионального роста. При подсыхании из листьев исчезает крахмал. Усиленный распад этого полисахарида не сопровождается накоплением гексоз, поскольку они используются в процессе дыхания. При потере воды усиливается распад белковых веществ.
Опыты с растениями тыквы и огурца (В.Н. Жолкевич, Т.Ф. Корецкая) показали, что почвенная засуха вызывает глубокое нарушение фосфорного обмена в корневой системе, что приводит к уменьшению в корнях количества АТФ, нуклеиновых кислот - РНК и ДНК, уменьшению содержания белков и увеличению в 7-10 раз сахаров, особенно фруктозы и глюкозы. Задержка фосфорилирования Сахаров вызывает уменьшение содержания органических кислот - пировиноградной кислоты, а также кислот цикла Кребса - б-кетоглютаровой, янтарной, фумаровой, щавелевоуксусной, яблочной и лимонной, являющихся акцепторами аммиака при синтезе аминокислот.
В корнях и в пасоке повышается содержание амидов, преимущественно глютамина. Следовательно, симптомами угнетения растений почвенной засухой прежде всего является нарушение энергетического обмена через уменьшение фосфорилирования, а также задержку синтеза белков. Длительные засухи приводят к резкому снижению урожая.
Как показали исследования Н.М. Сисакяна, при обезвоживании растительных тканей изменяется направленность действия ферментов в сторону гидролиза и задерживаются синтетические процессы и рост. Ростовые процессы задерживаются некоторое время даже после возобновления водоснабжения, поскольку восстановление синтетической направленности ферментов происходит не сразу. Различные части растения неодинаково устойчивы к засухе и неодинаково на нее реагируют.
Установлено, что растения, перенесшие небольшую засуху, повторную переносят с меньшими потерями. Однако устраивать искусственную закалку взрослых растений в производственных условиях трудно. П.А. Генкель предложил подвергать закаливанию наклюнувшиеся семена. Опыт показал, что растения, выросшие из семян, подвергнутых предпосевной закалке, приобретают повышенную устойчивость к засухе.
Растворы сахаров (глюкоза, галактоза, сахароза, маннит, лактоза, мальтоза, раффиноза) при инфильтрации в ткани листьев значительно повышают их устойчивость к высоким температурам. Считают, что сахара «консервируют» структуру митохондрии, которая становится нечувствительной к тепловому влиянию, и этим сохраняют функцию митохондрий - процесс дыхания и окислительное фосфорилирование (Ю.Г. Молотковский, И.М. Жесткова).
Засухоустойчивые сорта при значительном водном дефиците характеризуются синтетической направленностью в работе своих ферментов, тогда как у менее засухоустойчивых преобладает гидролитическая направленность. Засухоустойчивые сорта озимой пшеницы характеризуются большим содержанием связанной воды, которая трудно обменивается во время засухи, повышенной концентрацией клеточного сока в период цветения и налива зерна, высшим порогом коагуляции белков, более интенсивным накоплением сухого вещества зерна, более стойкой к неблагоприятным условиям пигментной системой. Этим и характеризуется физиологическая природа засухоустойчивости растений.
75. Фотосинтез в различных лучах спектра. Спектры поглощения хлорофиллов, каратиноидов, хлоропластов листа
Современные научные представления о фотохимических реакциях, входящих в процесс фотосинтеза, основываются на квантовой теории света. Согласно этой теории, каждая молекула вещества при фотохимических реакциях, т. е. при превращении вещества под влиянием света, поглощает частицу его -- фотон, или квант, который имеет достаточный запас энергии для того, чтобы вызвать в молекуле соответствующие изменения. Основное действие света определяется энергией как отдельных фотонов, так и световых потоков.