ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 19.10.2024
Просмотров: 24
Скачиваний: 0
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
СОДЕРЖАНИЕ
12. Химический состав и строение клеточной стенки, ее функциональное значение
34. Роль активной деятельности цитоплазмы и поглощении воды клеткой
54. Нарушение водообмена, его причины и последствия
89. Связь дыхания и брожения. Пути окисления пировиноградной кислоты в растительных тканях
95. Дегидрогеназы, их химическая природа и характер действия
130. Ингибиторы их физиологическая роль и применение в практике
176. Физиологические особенности засухоустойчивых сельскохозяйственных условий
Если квант меньше, чем необходимо энергии для данной реакции, то превращения молекулы не произойдет; а если он слишком велик, то превращение осуществляется, но расход энергии будет непродуктивным. В процессе фотосинтеза есть ряд промежуточных реакций, которые осуществляются до момента образования углевода. На каждую из этих реакций также расходуется определенное количество энергии. Продуктивность фотохимического процесса определяется количеством квантов, а не величиной отдельного кванта.
Энергия света Е, переносимая фотоном или квантом hv, равна:
Е=hv=hc/л
где h -- световая константа Планка, равняется 6,626*10-47 эрг/с, или 6,26196*10-84 Дж/с;
v -- частота колебаний данной световой волны;
л -- длина волны;
с -- скорость света в вакууме (примерно 3*108 м/c).
Согласно приведенному уравнению, квант света обладает энергией, пропорциональной его частоте и обратно пропорциональной длине волны.
В фотохимии употребляется единица количества света -- эйнштейн (е). Она равна 6,023> 1023 квантов монохроматического света. При поглощёнии -системой, способной к фотохимическим превращениям, одного е происходит фотохимическое превращение 6,023*1023 молекул, или одного моля вещества. Энергия одного е равна 6,023*1023 hv квантов или фотонов, и называют ее «молем» квантов.
В таблице приведены характеристики для ультрафиолетовых (видимых) и инфракрасных лучей.
Как установлено О. Варбургом и другими учеными, квантовый выход фотосинтеза, т. е. количество молекул реагирующего углекислого газа на квант света, который поглощается при фотосинтезе, равен приблизительно 0,25. Следовательно, на восстановление одной молекулы CО2 расходуется 4 кванта красного света:
ц - квантовый выход <1
Однако многочисленные экспериментальные определения квантовых выходов фотосинтеза показали, что при восстановлении одной молекулы СО2 поглощается 8--12 квантов света. Поглощение 4 квантов красных лучей может дать достаточное и даже избыточное количество энергии, а именно: 4 кванта красного света дают 502--669 кДж/моль, а для восстановления 1 моля, СО2 необходимо 470 кДж.
Очевидно, из 12 квантов энергия 4 квантов превращается в химическую энергию и связывается в продуктах фотосинтеза, энергия остальных 8 квантов используется на образование каких-то неустойчивых, высокоэнергетических продуктов, которые в дальнейшем, подвергаясь экзэргоническцм превращениям, отдают энергию на поддержание необходимых для фотосинтеза градиентов потенциалов; потом эта энергия деградирует в тепло.
Обратная квантовому выходу величина -- квантовые расходы
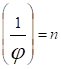
показывает, сколько квантов света должно быть поглощено растением для поглощения и превращения одной молекулы СО2 или выделения молекулы О2. При очень низкой интенсивности света расходы на фотосинтез не превышают 4 квантов. При увеличении интенсивности света количество Квантов на восстановление одной молекулы СО2 резко возрастает. При 12 квантах коэффициент полезного действия энергии активного света, который поглощается фотосинтетически, может составлять 19%, или 8--10% энергии общей радиации солнца, падающей на лист. Считают, что листья зеленых растений поглощают обычно 80--1 85% энергии видимой части солнечного спектра, пропускают 5--10%. и отражают около 10%.

89. Связь дыхания и брожения. Пути окисления пировиноградной кислоты в растительных тканях
Брожение -- это внутренний окислительно-восстановительный процесс, при котором акцептором электронов служит органическая молекула и суммарная степень окисления образующихся продуктов, отличается от степени окисления сбраживаемого вещества. Дыхание - это окислительный распад органических веществ при участии кислорода с образованием воды, углекислого газа и макроэргических соединений, которые используются клетками.
С.П.Костычев выдвинул положение о генетической связи процессов брожения и дыхания. При этом он опирался на следующие факты:
1. У высших растений был найден весь набор ферментов, который катализирует отдельные этапы процесса брожения.
2. При временном попадании в условия анаэробиоза высшие растения определенное время существуют за счет энергии, выделяющейся в процессе брожения. Правда, поскольку процесс брожения энергетически значительно менее эффективен, в анаэробных условиях рост растений приостанавливается. Кроме того, продукты брожения, в частности спирт, ядовиты, и их накопление приводит к гибели растения.
3. При добавлении к клеткам факультативных анаэробов (дрожжи) полусброженных Сахаров интенсивность дыхания у них резко возрастает, следовательно, полусброженные продукты являются лучшим субстратом дыхания по сравнению с неизмененными сахарами.
В настоящее время общепризнано, что первые этапы (гликолиз) протекают одинаково при процессах, как дыхания, так и брожения. Поворотным моментом является образование пировиноградной кислоты. В аэробных условиях пировиноградная кислота распадается до С02 и воды в результате декарбоксилирования и цикла Кребса (дыхание), тогда как в анаэробных она преобразуется в различные органические соединения (брожение). Организм обладает способностью при изменении условий переключать процессы, прекращая брожение и усиливая дыхание и наоборот.
Впервые в опытах Пастера было показано, что в присутствии кислорода процесс брожения у дрожжей тормозится и заменяется процессом дыхания. Одновременно резко сокращается распад глюкозы. Дело в том, что для брожения необходим НАДН, который в аэробных условиях окисляется. Это явление оказалось характерным для всех факультативных анаэробных организмов, включая высшие растения, и получило название эффекта Пастера.
Сокращение расхода глюкозы в присутствии кислорода целесообразно, поскольку при дыхательном распаде выход энергии значительно выше, а, следовательно, глюкоза используется более экономно. Однако осуществление разбираемого эффекта требует специальных механизмов. В зависимости от получаемого продукта различают разные типы брожения. При спиртовом брожении пировиноградная кислота, образовавшаяся в процессе гликолиза, декарбоксилируется с образованием уксусного альдегида при участии фермента пируватдекарбоксилазы, а затем восстанавливается до этилового спирта ферментом алкогольдегидрогеназой. Обе эти реакции не сопровождаются образованием АТФ. В связи с этим выход АТФ при спиртовом брожении такой же, как при гликолизе (первой фазы брожения и дыхания), и составляет две молекулы при распаде 1 моль глюкозы. Восстановленные никотинамидные коферменты НАДН + Н+, образовавшиеся в процессе гликолиза, не поступают в дыхательную цепь (у анаэробных организмов ее и нет), а используются для восстановления уксусного альдегида до спирта. Следовательно, энергетический выход процессов брожения крайне низок. Разные микроорганизмы осуществляют и разные типы брожения. Так, молочнокислые бактерии накапливают молочную кислоту. При этом пировиноградная кислота восстанавливается до молочной кислоты.
Для некоторых облигатных анаэробных микроорганизмов, например, азотфиксирующей бактерии Clostridiumpasteurianum, характерно образование в процессе брожения масляной кислоты, С02 и N2. Процесс брожения служит источником энергии для облигатных (обязательных) или факультативных анаэробных организмов.
95. Дегидрогеназы, их химическая природа и характер действия
Окисление органических веществ в живых тканях растений и животных происходит с участием ферментов: дегидрогеназ - активаторов и переносчиков водорода (электрона) дыхательного материала и оксидаз - активаторов молекулярного кислорода.
Дегидрогеназы (от де... и новолат. hуdrogenium - водород), ферменты, катализирующие отщепление водорода от органических веществ. Коферментами Д. обычно являются динуклеотиды:
- никотинамидадениндинуклеотид (НАД),
- никотинамидадениндинуклеотидфосфат (НАДФ) или флавинадениндинуклеотид (ФАД)
- флавинмононуклеотид (ФМН), которые акцептируют водород окисляемого вещества. Д. осуществляют первый этап биологического окисления.
Они играют большую роль в цикле Кребса, в гликолизе и в пентозофосфатном цикле. Некоторые Д., не связанные с коферментами, катализируют реакции окисления веществ непосредственно кислородом. Большая часть Д. в составе активного центра содержит металлы - цинк, марганец.
Дегидрогеназы действуют на протяжении всего процесса дыхания и по характеру действия делятся на аэробные и анаэробные. Первые переносят водород непосредственно на молекулярный кислород, а вторые - на какой-либо акцептор водорода. Дегидрогеназы относятся к двухкомпонентным ферментам - протеидам с пиридиновыми или флавиновыми (аллоксазиновыми) коферментами.
Пиридиновые дегидрогеназы относятся к группе анаэробных дегидрогеназ, которые являются акцепторами водорода субстрата. Специфичность их действия заключается в способности гидрирования и дегидрирования пиридиновых ядер.
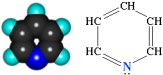
Пиридин
К флавопротеидным дегидрогеназам относятся ферменты, в состав которых фосфорный эфир рибофлавина.
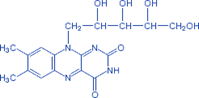
Рибофлавин