Файл: Липиды 1 Классификация липидов. Переваривание и всасывание липидов. Введение.doc
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 17.03.2024
Просмотров: 143
Скачиваний: 0
СОДЕРЖАНИЕ
По функциональной нагрузке в организме различают две группы липидов:
По химическому строению липиды разделяют на омыляемые и неомыляемые
Жирные кислоты - это алифатические карбоновые кислоты.
Функции ацилглицеролов в организме многообразны
Сложные липиды - главные компоненты биологических мембран
Фосфолипиды - сложные липиды, содержащие фосфор
Сфингофосфолипиды содержат в своем составе сфингозин
Плазмалогены - это эфирные производные глицерофосфолипидов
Гликолипиды - это сфинголипиды, содержащие углеводы
Неомыляемые липиды не гидролизуются щелочью
Высшие углеводороды - производные изопрена
Простагландины - продукты окисления жирных кислот
Липиды транспортируются в крови в составе липопротеинов
Номенклатура и характеристика липопротеинов
Липиды, поступившие из кишечника (экзогенные), транспортируются в кровотоке в составе хиломикронов
Переваривание липидов катализирует липаза
Так как липиды, в основном, нерастворимы в воде, то они подвергаются действию гидролитических ферментов только на границе раздела между липидами и водной фазой. Основным ферментом, который катализирует расщепление липидов пищи, является панкреатическая липаза. Этот фермент присутствует в соке поджелудочной железы и активируется в просвете кишечника. Дело в том, что в водной среде доступ к активному центру липазы заблокирован -спиральным участком её молекулы. На границе раздела жир-вода активный центр становится доступен для субстратов. Этому способствует также специальный белок, названный колипазой. Колипаза связывается с С-концом некаталитического участка молекулы панкреатической липазы. Происходящее вслед за этим изменение конформации обусловливает прикрепление ферментативного комплекса к липидной поверхности эмульсионных частиц. Активный фермент ускоряет реакцию гидролиза ацилглицеролов. Гидролиз триацилглицеролов сначала происходит в положении 1 или 3, что приводит к образованию диацилглицеролов, которые затем гидролизуются до 2-моноацилглицеролов. Меньшая часть (40%) моноацилглицеролов подвергается дальнейшему гидролизу до глицерола. Для остальной части процесс ферментативного гидролиза завершается на этапе образования 2-моноацилглицеролов. Необходимо отметить, что в расщеплении 2-моноацилглицеролов участвует также кишечная липаза, но активность этого фермента невысока.
В соке поджелудочной железы присутствуют и другие ферменты, способные расщеплять липиды. В частности, эстеразы катализируют преимущественно гидролиз эфиров жирных кислот с короткой цепью. В поджелудочной железе синтезируется профосфолипаза А2. Фермент приобретает активность только после воздействия в просвете кишечника трипсина, приводящего к отщеплению от него гептапептида. Фосфолипаза А2 катализирует отщепление молекулы жирной кислоты от фосфатидилхолина с образованием лизофосфатидилхолина .

Специфичность фосфолипаз
Продукты гидролиза липидов участвуют в образовании мицелл
Продукты ферментативного гидролиза жира взаимодействуют с водной средой, объединяются в мельчайшие частицы - мицеллы. По размеру они гораздо меньше, чем эмульсионные. Снаружи мицеллы, подобно эмульсионным частицам, покрыты слоем желчных кислот.
Основная часть всосавшихся в тонком кишечнике липидов принимает участие в ресинтезе триацилглицеролов.
Основная часть мицелл целиком или после предварительного (пристеночного) разрушения, всасываются через стенку тонкого кишечника. Желчные кислоты проходят через
барьер слизистой оболочки в связанном с липидами состоянии. В дальнейшем по венам кишечника они поступают в портальный кровоток. Оттуда они извлекаются печенью и затем снова поступают с желчью в двенадцатиперстную кишку. Всосавшиеся продукты расщепления липидов в клетках слизистой оболочки кишечника подвергаются процессам ресинтеза. Там имеются ферментные системы, которые могут превращать свободные жирные кислоты, моно- и диацилглицеролы в триацилглицеролы, фосфолипиды и эфиры холестерола.
Всосавшиеся ЖК активируются. Активация их заключается в присоединении остатка жирной кислоты к коферменту А с образованием ацил
КоА. Это происходит в гладком эндоплазматическом ретикулуме
1.
В клетках слизистой тонкого кишечника функционируют два пути ресинтеза триацилглицеролов. Это обусловлено поступлением туда при всасывании большого количества 2-моноацилглицеролов. Исключением, имеющим клиническое значение, является тот факт, триацилглицеролы, содержащие коротко- и среднецепочечные жирные кислоты (С6-С10), могут подвергаться всасыванию без предварительного расщепления ферментами. Из клеток слизистой тонкого кишечника они также попадают сразу в кровоток системы воротной вены. Этот механизм принципиально отличается от механизма всасывания и попадания в кровь основной массы липидов пищи, в составе которых содержатся жирные кислоты с количеством углеродных атомов более 10. Поэтому ряд полусинтетических лекарственных препаратов, приготовленных на базе кокосового масла и содержащих смесь триацилглицеролов с
коротко- и среднецепочечными жирными кислотами, эффективно используются при лечении заболеваний пищеварительной системы (синдромы малабсорбции вследствие недостаточности функции поджелудочной железы, непроходимость желчных путей, резекция тонкого кишечника, хронические заболевания печени, саркома брыжейки, непроходимость лимфатических сосудов кишечника, тропический спру, абеталипопротеинемия).

Липиды транспортируются в крови в составе липопротеинов
Новосинтезированные триацилглицеролы, фосфолипиды и другие всосавшиеся липиды покидают клетки слизистой, попадая сначала в лимфу, а с током лимфы - в кровь. В связи с тем, что большинство липидов, как уже отмечалось, нерастворимо в водной среде, транспорт их в лимфе, а затем - в плазме крови, осуществляется не так, как транспорт водорастворимых молекул подобного размера. Практически все липиды транспортируются в составе специальных частиц - липопротеинов.
Структура липопротеинов. Электронная микроскопия выделенных видов липопротеиновых частиц показала, что они представляют собой сферические частицы, диаметр которых уменьшается с увеличением плотности. Липопротеины состоят из ядра, включающего гидрофобные липиды - триацилглицеролы, эфиры холестерола , то время как наружная часть, находящаяся в контакте с плазмой крови, содержит амфифильные липиды: фосфолипиды, свободный холестерол. Белковые компоненты (апопротеины) своими гидрофобными участками располагаются во внутренней части липопротеинвых частиц, а гидрофильными - преимущественно, на поверхности.
Номенклатура и характеристика липопротеинов
Применение специального метода высокоскоростного центрифугирования (ультрацентрифугирования) в градиенте плотности некоторых солей позволяет разделить основные виды липопротеинов крови. Вследствие различной плотности из-за разного их химического состава, липопротеиновые частицы в ходе ультрацентрифугирования распределяются по определенным зонам в пробирке. Различают следующие классы липопротеиновых частиц: 1) хиломикроны - d<1,00г/мл; 2) липопротеины очень низкой плотности - d<1,006г/мл; 3) липопротеины промежуточной плотности - d=1,006-1,020г/мл; 4) липопротеины низкой плотности - d=1,02-1,063г/мл; 5) липопротеины высокой плотности - d=1,063-1,210г/мл. В настоящее время употребляется также старая номенклатура липопротеинов, основанная на их электрофоретической миграции в агарозном геле. Согласно ей ЛПОНП называются пре -липопротеины, ЛПНП - -липопротеины, а ЛПВП - -липопротеины.
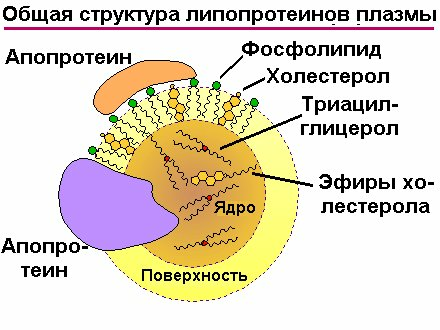 Рис.7.10. Все липопротеины плазмы крови являются частицами сферической формы, состоящими из ядра, включающего в себя триацилглицеролы и эфиры холестерола, окруженного тонким липидным монослоем молекул свободного холестерола и фосфолипидов. Аполипопротеины внедрены в поверхностную липидную оболочку таким образом, что их гидрофобные домены ориентированы в сторону ядра, а гидрофильные - наружу. Такая конфигурация является высоко устойчивой и облегчает растворимость микрокапель неполярных липидов. | Частица ЛПНП |
Характеристика липопротеинов плазмы крови
| Липопротеины | Источник | Диаметр нм | Плотность | % белка | % липиды | % от общего количества липидов | |
| | | | | | | ТАГ | ЭХ |
| Хиломикроны | Кишечник | 90 - 1000 | < 0,95 | 1 - 2 | 98 - 99 | 88 | 3 |
| ЛПОНП | печень | 30 - 90 | 0,95-1,006 | 7 - 10 | 90 - 93 | 56 | 15 |
| ЛППП | ЛПОНП | 25 - 30 | 1,006-1,009 | 11 | 89 | 29 | 34 |
| ЛПНП | ЛПОНП | 20 - 25 | 1,019-1,063 | 21 | 79 | 13 | 48 |
| ЛПВП2 | печень | 10 - 20 | 1,063-1,125 | 33 | 67 | 16 | 31 |
| ЛПВП3 | ? | 7,5 - 10 | 1,125-1,210 | 57 | 43 | 13 | 29 |