Файл: Липиды 1 Классификация липидов. Переваривание и всасывание липидов. Введение.doc
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 17.03.2024
Просмотров: 140
Скачиваний: 0
СОДЕРЖАНИЕ
По функциональной нагрузке в организме различают две группы липидов:
По химическому строению липиды разделяют на омыляемые и неомыляемые
Жирные кислоты - это алифатические карбоновые кислоты.
Функции ацилглицеролов в организме многообразны
Сложные липиды - главные компоненты биологических мембран
Фосфолипиды - сложные липиды, содержащие фосфор
Сфингофосфолипиды содержат в своем составе сфингозин
Плазмалогены - это эфирные производные глицерофосфолипидов
Гликолипиды - это сфинголипиды, содержащие углеводы
Неомыляемые липиды не гидролизуются щелочью
Высшие углеводороды - производные изопрена
Простагландины - продукты окисления жирных кислот
Липиды транспортируются в крови в составе липопротеинов
Номенклатура и характеристика липопротеинов
Липиды, поступившие из кишечника (экзогенные), транспортируются в кровотоке в составе хиломикронов
Производное фосфатидилинозита - соединение фосфатидилинозит-4,5-бисфосфат является важным компонентом биологических мембран. При стимуляции соответствующим гормоном он расщепляется. Продукты его расщепления (диацилглицерол и инозитолтрифосфат) служат в качестве внутриклеточных посредников действия гормонов.
 | 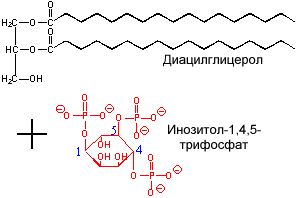 |
С глицерофосфолипидами метаболически очень тесно связаны лизофосфолипиды. В их составе содержится только один остаток жирной кислоты. Примером может служить лизофосфатидилхолин, который играет важную роль в метаболизме фосфолипидов.
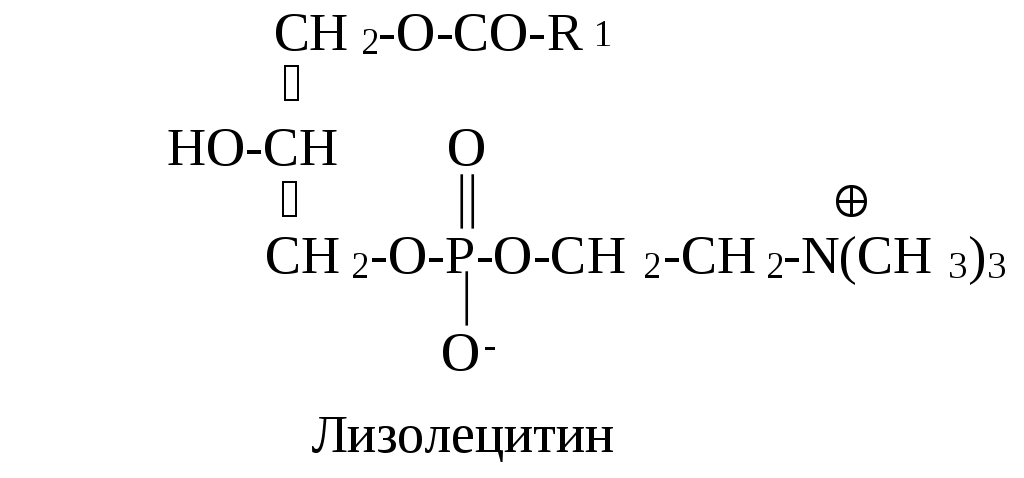 Лизофосфатидилхолин способен вызывать лизис эритроцитов. Поэтому наличие в змеином яде большого количества лизофосфолипидов является причиной гемолиза, который наблюдается при укусах змей.
Лизофосфатидилхолин способен вызывать лизис эритроцитов. Поэтому наличие в змеином яде большого количества лизофосфолипидов является причиной гемолиза, который наблюдается при укусах змей. Сфингофосфолипиды содержат в своем составе сфингозин


Сфингомиелин
Первая часть слова "сфинго" свидетельствует о том, что в состав молекулы входит вместо глицерина двухатомный ненасыщенный спирт - сфингозин. Наиболее широко распространенным в организме представителем этой группы соединений является сфингомиелин Сфингомиелин обнаружен в мембранах растительных и животных клеток; особенно богата сфингофосфолипидами нервная ткань, и в частности, мозг.
Характерной особенностью фосфолипидов является их дифильность, то есть способность растворяться как в водной среде, так и в нейтральных липидах. Это обусловлено наличием у фосфолипидов выраженных полярных свойств. При рН 7,0 их фосфатная группа всегда несет отрицательный заряд. Азотсодержащие группировки в составе фосфатидилхолина (холин) и фосфатидилэтаноламина (этаноламин) при рН 7,0 несут положительный заряд. Таким образом, при рН 7,0 эти глицерофосфолипиды представляют собой биполярные цвиттерионы и их суммарный заряд равен нулю. Остаток серина в молекуле фосфатидилсерина содержит -аминогруппу и карбоксильную группу. Следовательно, при рН 7,0 молекула фосфатидилсерина имеет две отрицательно и одну положительно заряженных группы и несет суммарный отрицательный заряд.
В то же время, радикалы жирных кислот в составе фосфолипидов не имеют электрического заряда в водной среде и таким образом обусловливают гидрофобность части молекулы фосфолипида. Наличие полярности за счет заряда полярных групп обусловливают гидрофильность. Поэтому на поверхности раздела масло-вода фосфолипиды располагаются таким образом, чтобы полярные группы находились в водной фазе, а неполярные группы - в масляной. За счет этого в водной среде они образуют бимолекулярный слой, а при достижении некоторой критической концентрации - мицеллы.]
На этом основано участие фосфолипидов в построении биологических мембран.
Обработка находящегося в водной среде дифильного липида ультразвуком приводит к образованию липосом. Липосома представляет собой замкнутый липидный бислой, внутри которого оказывается часть водной среды. Липосомы находят применение в клинике, косметологии в качестве

своеобразных контейнеров и переносчиков лекарств, питательных веществ к определенным органам и для комбинированного действия на кожу.
Функциональная роль фосфолипидов не ограничивается их участием в построении биомембран. Так, они являются регуляторами активности ферментов. К примеру, фосфатидилхолин, фосфатидилсерин, сфингомиелин активируют или ингибируют активность ферментов, катализирующих процессы свертывания крови. Регуляторная функция липидов заключается в том, что ряд гормонов (половые, гормоны коры надпочечников) являются производными липидов. Кроме того фосфолипиды
Выполняют детергентную функцию в кишечнике и желчном пузыре. Они являются важным структурным компонентом желчи, наряду со свободным холестеролом и с желчными кислотами. Изменение соотношения любого из этих компонентов приводит к осаждению и формированию желчных камней. Фосфолипиды - это также важный компонент смешанных мицелл, которые образуются в ходе переваривания липидов.
Является источником арахидоновой кислоты — предшественника эйкозаноидов
Являются источниками вторичных мессенджеров - диацилглицерола и инозитолтрифосфата, о чем уже упоминалось выше
Обеспечивают прикрепление белков к мембране. Некоторые внеклеточные белки прикрепляются к внешней стороне плазматической мембраны за счет образования ковалентных связей с фосфатидилинозитолом. Примером таких белков могут служить ферменты: щелочная фосфатаза, липопротеин липаза, холинэстераза.
Принимают участие в формировании транспортных форм других липидов
Могут выполнять энергетическую функцию
Явяляются компонентом сурфактанта легких (см. ниже)
Плазмалогены - это эфирные производные глицерофосфолипидов
Эти соединения содержат простую эфирную связь при С1 скелета глицерола.
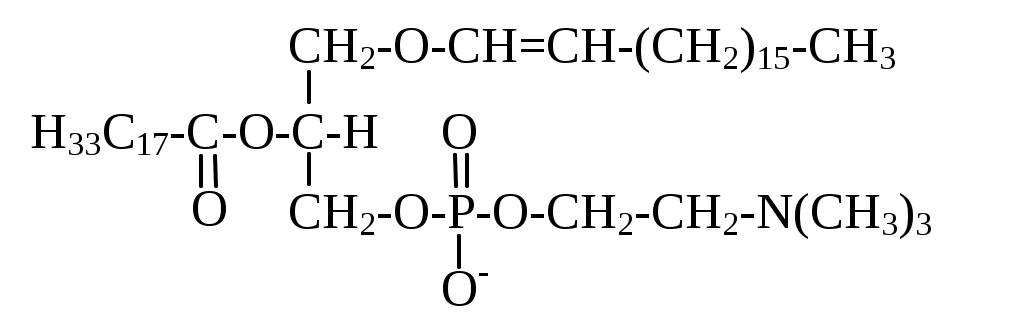
При полном их гидролизе наряду с глицеролом, фосфорной кислотой и спиртом (холином) образуется 1 молекула жирной кислоты и альдегид. В настоящее время известны три основных вида плазмалогенов: холин, этаноламин и серин плазмалогены. Этаноламиновые плазмалогены преобладают в составе миелина. Холиновые плазмалогены распространены в ткани сердца. К холиновым плазмалогенам, в частности, относится фактор активации тромбоцитов - медиатор, обладающий исключительной биологической активностью. Он способен вызывать клеточный ответ в концентрации 10
-11 М. В структуре его молекулы ко второму углеродному атому глицерола присоединен остаток уксусной кислоты.
Присутствие в положении С-2 ацетильной группы вместо длинноцепочечной жирной кислоты делает это соединение растворимым в воде в физиологических концентрациях. Синтезируясь в клетках эндотелия сосудистой стенки, фактор активации тромбоцитов содержится в тромбоцитах и регулирует сосудистый тонус, способствует адгезии лейкоцитов, является прокоагулянтным фактором. В базофилах, нейтрофилах, эозинофилах, макрофагах и моноцитах он синтезируется в ответ на образование на их поверхности комплексов антигена с иммуноглобулином Е. Высвобождаясь из этих клеток, фактор активации тромбоцитов функционирует как медиатор повышенной чувствительности, воспалительных реакций и анафилактического шока. Он вызывает ответную реакцию в печени, сердце, гладкой мускулатуре, в матке и легочной ткани.
Гликолипиды - это сфинголипиды, содержащие углеводы
Гликолипиды широко представлены в тканях. Особенно богаты ими миелиновые оболочки нервов. В состав гликолипидов также входит спирт - сфингозин. Гликолипиды не содержат фосфорной кислоты. Молекулы их имеют полярные, гидрофильные углеводные группы, чаще всего D-галактозу. Различают две группы гликолипидов - цереброзиды и ганглиозиды.В состав молекулы цереброзида входит сфингозин, связанный сложноэфирной связью с остатком жирной кислоты (этот комплекс называется церамид).
Обнаруженные в цереброзидах жирные кислоты необычны в том отношении, что они содержат 24 атома углерода. Наиболее часто встречаются нервоновая, цереброновая и лигноцериновая кислоты. Углеводная часть цереброзида представлена D-галактозой, которая присоединена к сфингозину. В состав цереброзидов различных тканей организма, за исключением нервной, может входить глюкоза вместо галактозы. Ганглиозиды имеют более сложное строение. В состав молекулы ганглиозидов, помимо сфингозина, входит олигосахарид, содержащий остатки глюкозы и галактозы, а также одну или несколько молекул сиаловых кислот. Сиаловые кислоты - это производные аминосахаров. Доминирующими в составе ганглиозидов являются N-ацетил-D-глюкозамин и N-Ацетилнейраминовая кислота.
Г
Функции гликолипидов в организме
| 1. Опосредуют | Межклеточное взаимодействие Взаимодействие клеток с межклеточным матриксом Взаимодействие клеток с микроорганизмами |
| 2. Модулируют | Пролиферацию клеток, угнетая её (апоптоз, нарушение клеточного цикла) Активность протеинкиназы Активность рецептора к фактору роста |
| 3. Поддерживают | Структурную прочность мембран Конформацию мембранных белков |