Файл: 1. Современные представления о строении и функции мембран.docx
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 10.04.2024
Просмотров: 236
Скачиваний: 0
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.

Результатом взаимодействия медиаторов с ионотропными рецепторами хемозависимых ионных каналов постсинаптической мембраны может быть ее кратковременная гиперполяризация. В результате понижается возбуди¬мость постсинаптического нейрона и в нем не происходит генерация по¬тенциала действия. Гиперполяризация постсинаптической мембраны в об¬ласти одного синапса называется тормозным постсинаптическим потен¬циалом (ТПСП), такие химические синапсы называются тормозными. По аналогии медиаторы, вызывающие ТПСП, называются тормозными медиа¬торами. Взаимодействие тормозного медиатора с рецепторами постсинап¬тической мембраны открывает хлорные и калиевые ионные каналы, что резко повышает ионную проводимость постсинаптической мембраны для ионов хлора (1а) или калия (/*). В соответствии с электрохимическими градиентами ионов калия и хлора по обе стороны от постсинаптической мембраны, ТПСП представляет собой сдвиг потенциала на постсинаптиче¬ской мембране в сторону увеличения его величины. В результате происхо¬дит гиперполяризация постсинаптической мембраны и увеличение порога ее возбудимости.
При передаче возбуждения через аксосоматические синапсы на мембра¬не одного нейрона одновременно возникают ВПСП и ТПСП. При этом происходит сложение (суммация) амплитуды ВПСП и ТПСП многих си¬напсов. Если в условиях суммации ВПСП и ТПСП суммарная амплитуда ВПСП достигает критического уровня деполяризации, то в мембране ак¬сонного холмика нейрона электротонически открываются потенциалзави¬симые натриевые ионные каналы и генерируется потенциал действия. В результате возбуждение передается от нервных терминалей пресинаптиче¬ского нейрона на постсинаптический нейрон.
15. Молекулярные механизмы сокращения и расслабления скелетной мышцы. Электромеханическое сопряжение.
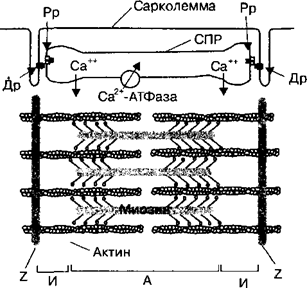 Во время укорочения мышцы уменьшается длина саркомеров. При этом филаменты актина и миозина, входящие в состав саркомеров, не изменяют своей длины. Уменьшение длины саркомеров происходит в результате движения (скольжения) тонких филаментов с двух сторон саркомера к его
Во время укорочения мышцы уменьшается длина саркомеров. При этом филаменты актина и миозина, входящие в состав саркомеров, не изменяют своей длины. Уменьшение длины саркомеров происходит в результате движения (скольжения) тонких филаментов с двух сторон саркомера к его центру относительно толстых миозиновых филаментов. Цикл поперечного мостика во время сокращения мышцы - это связывание, укорочение, высвобождение и вновь связывание поперечного мостика миозина с актиновым филаментом.
В состоянии покоя поперечный мостик расположен параллельно мио- зиновому стержню, а его головка — перпедикулярно относитель¬но последнего, и не прикреплена к актиновому филаменту (стадия 1). По¬сле того, как ионы Са2+ высвобождаются из саркоплазматического ретику¬лума (описано ниже) и активируют сокращение мышцы, активновый фи- ламент может связываться с миозиновой головкой (стадия 2). После свя¬зывания головка наклоняется (стадия 3), используя энергию, заключенную в миозин-АТФазном комплексе головки. Это является основной силой по¬перечного мостика, которая растягивает его эластический компонент для того, чтобы произвести энергию движения вдоль активного филамента (стадия 4). В результате головка мостика совершает движение на 40° к цен¬тру саркомера и перемещает при этом на 10 нм нить актина, В это время от головки высвобождаются АДФ и монофосфат, что необходимо для за-хвата головкой миозина новой молекулы АТФ. Только после этого головка
миозина отделяется от молекулы актина, необходимой для размыкания по-перечного мостика с актином (положение 5).
Цикл будет продолжаться до тех пор, пока концентрация ионов Са++ в межфибриллярном пространстве не снижается до подпороговой величины, что приведет к расслаблению мышцы, или пока не будут ис¬черпаны запасы АТФ. В последнем случае в мышце возникает спазм или мышечная контрактура (rigor), при которой в течение нескольких секунд или минут отсутствует расслабление мышцы. Таким образом, функция АТФ проявляется в двух точках процесса со¬кращения мышцы. Во-первых, АТФ является источником энергии для со¬кращения. Во-вторых, АТФ отсоединяет поперечной мостик для того, что¬бы повторялся цикл поперечных мостиков «замыкание—сокращение—раз¬мыкание».
Расслабление скелетной мышцы
Расслабление скелетной мышцы начинается только после того, как кон¬центрация ионов Са2+ в саркоплазме мышечных клеток снижается ниже порогового уровня, т. е. ниже 10 мкМ/л, или 10"5 М. Расслабление мышцы обусловливают три процесса:
1. Отсоединение (диссоциация) ионов Са2+ от субъединицы С тропонина.
2. Диффузия ионов Са2+ в сторону сарко¬плазматического ретикулума.
3. Активный транспорт ионов Са2+ внутрь СПР при участии Са2+-АТФазы.
В начальный момент расслабления длина мышцы остается постоянной. Это происходит из-за того, что первоначально происходит расслабление саркомеров, находящихся на концах мышц, в то время как центральные саркомеры остаются укороченными. Это состояние характерно лишь для начального момента мышечного расслабления, а именно фазы изометрического расслабления. Процесс расслабления во всех саркомерах усиливается после того как сила сокращения мышцы уменьшается примерно на 20— 30 %. Это обусловлено тем, что во время расслабления мышцы уменьшает¬ся подвижность тропомиозина по поверхности белковых цепей актина примерно с той же скоростью, что и сила сокращения. В

результате чем меньше сила сокращения мышцы, тем меньше открыто мест связывания на актиновых филаментах для поперечных мостиков миозина.
Электромеханическое сопряжение - процесс, при котором потенциалы действия, генерируемые на сарколемме в области нервно-мышечного синапса, активируют сокращение мышцы.
В состоянии покоя мышечного волокна Са2+-АТФаза саркоплазматиче- ского ретикулума активно закачивает ионы Са2+ из саркоплазмы в его цис¬терны, где большинство ионов находится в обратимо связанном состоянии с белком кальсеквестрином. Для того чтобы вызвать сокращение, потенциал действия мышцы, вы- зваемый двигательным нервом, распространяется по сарколемме мышеч¬ной клетки и внутрь поперечных трубочек (t-система), открывая кальцие¬вые ионные каналы мембраны.
Далее происходит выход ионов Са2+ в саркоплазму. Когда концентрация ионов Са2+ в межфибриллярном пространстве, т. е. в саркоплазме между нитями актина и миозина, увеличивается до 10 мкМ/л (1(Г5 М) и более, ионы Са2+ начинают связываться с тропонином С. Активация тропонина С при связывании с ионами Са2+ вызывает смещение тропонин-тропомиози- нового комплекса примерно на 25° вокруг поверхности актинового фила¬мента и погружение молекул тропомиозина вглубь между белковыми цепя¬ми актина. В результате на актиновых филаментах открываются места свя¬зывания для головок поперечных мостиков миозина и начинается цикл их сокращения. При этом начинается сокращение всей скелетной мышцы, которое будет продолжаться до тех пор, пока концентрация ионов Са2+ в саркоплазме не станет пороговой или сверхпороговой для поддержания со¬кратительного процесса.
16. Виды и режимы сокращения мышц. Тетанус, виды тетануса, условия их возникновения.
1. Быстрое сокращение и расслабление скелетной мышцы в лабораторных условиях в ответ на стимуляцию называется одиночным мышечным сокращением (ОМС) – 1ый вид. Суммация ОМС при увеличении частоты раздражения приводит к тетаническому сокращению – 2ой вид.
2. Сокращение отдельного мышечного волокна подчиняется закону «все или ничего».
3. Амплитуда сокращения целой мышцы не подчиняется закону «все или ничего» и может увеличиваться при увеличении частоты и силы раздражения.
4. Амплитуда сокращения также увеличивается при увеличении количества мышечных волокон, участвующих в сокращении. При сокращении всех мышечных волокон амплитуда сокращения максимальная (оптимум). При дальнейшем увеличении частоты и силы раздражения амплитуда сокращения снижается – пессимум.
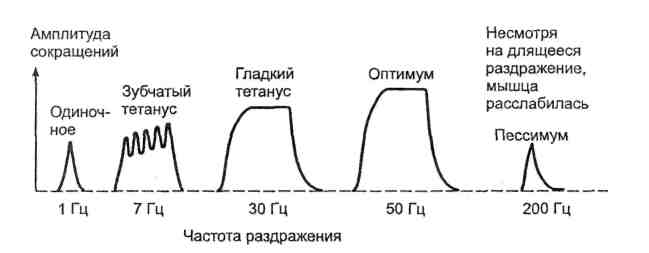 5. При невысокой частоте стимуляции каждое последующее раздражение попадает в фазу расслабления ОМС, и поэтому суммация ОМС будет неполной - зубчатый тетанус. При более высокой частоте стимуляции каждое последующее раздражение попадает в фазу сокращения ОМС, мышца не успевает расслабиться - гладкий тетанус.
5. При невысокой частоте стимуляции каждое последующее раздражение попадает в фазу расслабления ОМС, и поэтому суммация ОМС будет неполной - зубчатый тетанус. При более высокой частоте стимуляции каждое последующее раздражение попадает в фазу сокращения ОМС, мышца не успевает расслабиться - гладкий тетанус. 6. Режимы сокращения мышцы. Сокращение называется изометрическим (сопротивление давлению), если напряжение мышцы растет, но укорочения не происходит. Если мышца укорачивается при неизменном напряжении, то такое сокращение называется изотоническим (поднятие груза). Ауксотоническое – сокращение, при котором меняется и напряжение, и длина мышцы.
17. Утомление скелетной мышцы, его признаки. Механизмы мышечного утомления.
Утомлением называется временное снижение способности мышцы разви¬вать усилие в результате предшествующей физической активности. При этом происходит снижение всех физиологических параметров сокращения: скорости нарастания и снижения силы мышцы, скорости укорочения мышцы, работы и мощности.
Признаки утомления:
- увеличение латентного периода
- снижение амплитуды сокращения
- снижение скорости укорочения и расслабления
- Неполное расслабление ( остаточные контрактуры)
- Субъективные ощущения
По механизму возникновения утомления различают центральный и пе-риферический процессы. Центральное утомление представляет собой про¬цессы, происходящие во время мышечной работы на различных уровнях нервной регуляции движения, однако среди причин утомления оно состав¬ляет не более 10 %. При этом показана относительная неутомляемость нервных волокон при проведении по ним потенциалов действия. Перифе¬рическое утомление обусловлено процессами, происходящими на уровне нервно-мышечного синапса t-системы мышечных клеток, и ему принадле¬жит основная роль в снижении силы, скорости укорочения и расслабле¬ния, а также работу и мощности сокращающихся мышц. Снижение возбудимости сарколеммы является основной причиной в механизме периферического утомления. Основными факторами, снижающими активность Na+/K+-Hacoca, являются изменение концентрационного гра¬диента ионов Na+ и К+ по обе стороны мембраны мышечного волокна в области t-системы сарколеммы в результате генерации потенциалов дейст¬вия на мембране. Одновременно происходит снижение концентрации ионов Na+ на внеш¬ней поверхности мембраны мышечной клетки и повышение с внутренней стороны. Это приводит к деполяризации сарколеммы до величины мем¬бранного потенциала, равной в среднем — 60 мВ.
Во время физической работы у человека в плазме крови повышается концентрация адреналина и норадреналина. Эти гормоны стимулируют ра¬боту Na+/K+-Hacoca.
18. Гладкие мышцы. Типы гладких мышц. Молекулярные механизмы сокращения и расслабления гладкой мышцы.
Гладкие мышцы – это веретенообразные одноядерные мышечные клетки, которые составляют стенку внутренних органов, сосудов и активируются автономной нервной системой.