Файл: Лобзин В. С., Решетников М. М. Аутогенная тренировка.doc
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 04.05.2024
Просмотров: 488
Скачиваний: 1
СОДЕРЖАНИЕ
Глава 1. ТЕОРЕТИЧЕСКИЕ И ПРАКТИЧЕСКИЕ ОСНОВЫ АУТОГЕННОЙ ТРЕНИРОВКИ
ОСНОВНЫЕ ИСТОЧНИКИ АУТОГЕННОЙ ТРЕНИРОВКИ
ИНДИЙСКАЯ СИСТЕМА ХАТХА-ЙОГА И РАДЖА-ЙОГА
АКТИВНАЯ РЕГУЛЯЦИЯ МЫШЕЧНОГО ТОНУСА
АУТОГЕННАЯ ТРЕНИРОВКА И СХОДНЫЕ С НЕЙ МЕТОДЫ
Глава 2. ОБОСНОВАНИЕ МЕТОДА И ЧАСТНЫХ ПРИЕМОВ АУТОГЕННОЙ ТРЕНИРОВКИ
НЕЙРОФИЗИОЛОГИЧЕСКИЕ И НЕЙРОПСИХОЛОГИЧЕСКИЕ МЕХАНИЗМЫ САМОРЕГУЛЯЦИИ
ОСОЗНАВАЕМАЯ И НЕОСОЗНАВАЕМАЯ ПСИХИЧЕСКАЯ АКТИВНОСТЬ
ТЕОРИЯ УСТАНОВКИ И ЕЕ КЛИНИКО-ПСИХОЛОГИЧЕСКИЕ АСПЕКТЫ
УЧЕНИЕ ОБ ОБЩЕМ АДАПТАЦИОННОМ СИНДРОМЕ
ФЕНОМЕН ПСИХОЛОГИЧЕСКОЙ ЗАЩИТЫ
НЕЙРОФИЗИОЛОГИЧЕСКИЕ ЭФФЕКТЫ ТРЕНИРОВАННОЙ МЫШЕЧНОЙ РЕЛАКСАЦИИ
НЕЙРОПСИХОЛОГИЧЕСКИЕ ЭФФЕКТЫ КОМПЛЕКСА УПРАЖНЕНИЙ АУТОГЕННОЙ ТРЕНИРОВКИ
Глава 3. КЛИНИЧЕСКИЕ ВАРИАНТЫ АУТОГЕННОЙ ТРЕНИРОВКИ
АУТОГЕННАЯ ТРЕНИРОВКА ПЕРВОЙ СТУПЕНИ (АТ-!)
АУТОГЕННАЯ ТРЕНИРОВКА ВЫСШИХ СТУПЕНЕЙ
Глава 4. ПРИМЕНЕНИЕ АУТОГЕННОЙ ТРЕНИРОВКИ У ЗДОРОВЫХ ЛЮДЕЙ
АУТОГЕННАЯ ТРЕНИРОВКА В СПОРТИВНОЙ ПРАКТИКЕ
ПРОФЕССИОНАЛЬНО-ПРИКЛАДНОЕ ПРИМЕНЕНИЕ АУТОГЕННОЙ ТРЕНИРОВКИ
Глава 5. ОБУЧАЮЩИЙ КУРС АУТОГЕННОЙ ТРЕНИРОВКИ
КОМПЛЕКТОВАНИЕ ГРУПП ДЛЯ ГЕТЕРОГЕННЫХ ТРЕНИРОВОК
ИНДИВИДУАЛЬНОЕ ПРИМЕНЕНИЕ АУТОГЕННОЙ ТРЕНИРОВКИ
МЕТОДИКА И ТЕХНИКА ГРУППОВЫХ ЗАНЯТИЙ
СОЧЕТАНИЕ АУТОГЕННОЙ ТРЕНИРОВКИ С МУЗЫКОТЕРАПИЕЙ
Глава 6. СПЕЦИАЛЬНЫЕ УПРАЖНЕНИЯ И МЕТОДЫ НА ОСНОВЕ АУТОГЕННОЙ ТРЕНИРОВКИ
МЕТОДЫ ИНДИВИДУАЛЬНОЙ ПСИХОГИГИЕНЫ И ПСИХОКОРРЕКЦИИ
НЕКОТОРЫЕ УЗКОСПЕЦИАЛИЗИРОВАННЫЕ ПРИЕМЫ
Глава 7. ПРИМЕНЕНИЕ АУТОГЕННОЙ ТРЕНИРОВКИ В КЛИНИЧЕСКОЙ ПРАКТИКЕ
ПОГРАНИЧНЫЕ СОСТОЯНИЯ И НЕВРОЗЫ
АУТОГЕННАЯ ТРЕНИРОВКА ПРИ СОМАТИЧЕСКИХ ЗАБОЛЕВАНИЯХ
Наши исследования электроэнцефалографических изменений у больных неврозами во время сеансов аутогенной тренировки показали, что по сравнению с фоновыми данными в первой фазе сеанса аутотренинга (релаксация) наблюдается замедление альфа-ритма и появление медленных тетаволн, преимущественно в передних отделах головного мозга (лобные, лобно-теменные и височные отведения). Во второй фазе, при переходе к активному самовнушению, мед-
ленных волн становится меньше, и основной фон составляет хорошо выраженный альфа-ритм (индекс альфа-ритма увеличивается). Такая фазность биоэлектрической активности прослеживалась у 75 % больных. Это позволило заключить, что первая фаза аутогенной тренировки проявляется снижением корковой активности и характеризуется преобладанием тормозных процессов в коре больших полушарий. Во второй фазе регистрируемые на ЭЭГ процессы свидетельствуют о концентрации внимания, состоянии бодрствования и активной корковой деятельности (чего не наблюдается при гипнотическом внушении). Нейропсихологические корреляты отмеченных явлений выражались в двух совершенно четко разграничиваемых больными состояниях. Вначале, во время мышечного расслабления, появляется ощущение тяжести во всем теле, которое затем сменяется чувством легкости, «невесомости», «парения в воздухе».
Проведенные А. И. Скрыпниковым (1982) на высоком методическом и техническом уровне исследования ЭЭГ-коррелятов различных фаз аутогенного состояния у здоровых людей показали, что в первую фазу, по сравнению с исходным уровнем, повышается активность бета-ритма, которая достигает наибольшей выраженности к 8 —14-й минуте, увеличиваясь на 100—140% по отношению к фону. Увеличение энергии альфа-ритма начинается примерно на 8-й минуте сеанса аутогенной тренировки и достигает максимума (150%) к 16-й минуте. Энергия медленной составляющей ЭЭГ (дельта- и тета-ритмы) в процессе сеанса снижается и после завершения аутотренинга составляет соответственно 37 и 86% от фонового уровня. Это позволило автору сделать заключение, что уже при выполнении релаксирующих упражнений достигается активирующее воздействие на ЦНС, усиливающееся при аутогенной стимуляции к концу сеанса.
Сходные данные были получены и в ряде других исследований. К. Шипошем (1980) было показано, что в процессе аутогенной тренировки выявляется тенденция к синхронизации ритмов в области доминантного полушария и относительная активация субдоминантного полушария, что не наблюдается во время дремоты или естественного сна. Эти данные еще раз указывают на качественные отличия аутогенного состояния от других фазовых состояний (сон, гипноз).
Релаксация оказывает нормализующее влияние на основной обмен и насыщение крови кислородом [Решетников М. М., 1981], обмен йода и холестерина (рис. 6). Содержание кортизона в сыворотке крови в состоянии релаксации несколько снижается [Рожнов
B. Е., 1979]. Плетизмографические исследования показали, что в состоянии релаксации при вызывании ощущений тепла объем циркулирующей крови увеличивается [Sapir M. et al., 1975], объективно констатируются изменения лейкоцитарной формулы и содержания сахара в крови. Результаты исследований
C. Drunkenmolle, M. Bartusch, которые приводит G. Klumbies (1983), показали, что во время «тепловых упражнений» наблюдаются уменьшение общего пери-
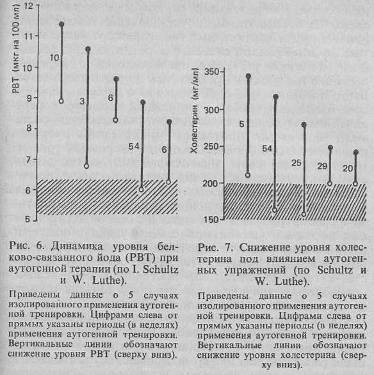
ферического сопротивления сосудистого русла и усиление кровообращения в правой руке в среднем у 62% пациентов (методом веноплетизмографии). Н. Prill, применявший аутогенную тренировку в гинекологической практике, отмечает нормализацию овариальных функций при дисменорее и вторичной аменорее. Под влиянием аутогенной тренировки снижается и уровень холестерина (рис. 7). Многочисленные физиологические эффекты релаксации, усиливающиеся при целенаправленном самовнушении, определяют широкий спектр показаний к применению метода в клинической практике.
За последние годы физиология и патофизиология нервной системы существенно пополнились сведениями о структурах и функциях глубинных отделов мозга. Успехи современной электроники и стереотаксической техники предоставили новые возможности для изучения и раскрытия структурно-функциональной организациицелого ряда образований центральной нервной системы. В частности, были установлены тесные функциональные и структурные связи неокортекса с гипоталамо-лимбической системой и ретикулярной формацией, а также уточнены представления о передаче нервного импульса от рецепторов до корковых клеток. Было показано, что одновременно с нервным процессом, имеющим залповый, линейный характер, с передачей импульсов по классическим лемнисковым системам, постоянно протекают циклические процессы с вовлечением нейронных «кругов» на разных уровнях регуляции за счет многочисленных коллатералей, связывающих специфические системы с экстралемни-сковыми (неспецифическими). Это сочетание залповой и циклической составляющих нервного процесса может рассматриваться как одно из основных проявлений корковых механизмов саморегуляции, обеспечивающих непрерывную адаптацию к быстро изменяющимся фазным раздражителям различной модальности, а также к длительной постоянной импульсации. Введенное вначале в кибернетике, понятие прямой и обратной связи нашло многочисленные подтверждения в нейрофизиологии. Было установлено существование постоянной «кольцевой» связи между афферен-тацией,

рецепцией и моторикой, а в самой рецепции были разграничены информационные, осуществляемые по лемнисковым системам и стресс-мобилизующие функции, связанные с экстралемнисковыми системами. Таким образом, в каждом движении присутствует в качестве обязательного элемента рецепция, а каждый рецепторный акт имеет и моторный компонент. С этой точки зрения, релаксация представляется как весьма специфический процесс, при котором существенно изменяются соотношения рецепции и моторики, а также отражение последних в сознании. Если в обычных условиях жизни и деятельности моторные акты осуществляются при минимуме рецепции (ее сознательного отражения), то в процессе систематических упражнений в мышечной релаксации целенаправленно тренируется рецептивный компонент при минимизации роли моторного.
Важным открытием современной нейрофизиологии явилось определение неспецифического активирующего влияния ретикулярной формации ствола мозга на кору больших полушарий (G. Moruzzi, Н. Magoun). В частности, в опытах на животных было показано, что при раздражении ретикулярной формации во время естественного сна (через вживленные электроды) возникает быстрое пробуждение. Одновременно на электроэнцефалограмме беспорядочные синхронизированные «сонные» колебания сменялись активным, десинхронизированным ритмом, также свидетельствующим о переходе к состоянию бодрствования. Эта реакция генерализованного пробуждения проявлялась при раздражении центральной части ретикулярной формации. При стимуляции ростральных отделов ретикулярной формации с неспецифическими ядрами таламуса и диффузной таламокортикальной системой у бодрствующего животного возникала реакция настороженности, обостренного восприятия и внимания (Д. Линдсли). Раздражение бульбарного отдела ретикулярной формации приводило к «глобальному» торможению или «облегчению» спинномозговых моторных разрядов. Одновременно были найдены корковые зоны (4S), стимуляция которых вызывала снижение мышечного тонуса, а также путь из зоны 4S к ретикулярной формации продолговатого мозга. Принимая все раздражения от экстеро- и интерорецеп-торов (при этом самым обширным рецептивным полем, как известно, является мышечная система), ре
тикулярная формация не только осуществляет их переработку, но и специфически активирует кору в зависимости от характера поступающих раздражений. Участвуя в процессах регуляции уровня бодрствования, ретикулярная формация имеет самые непосредственные связи с так называемым «висцеральным мозгом», интегрирующим раздражения от органов чувств и внутренней среды организма, которые в значительной степени определяют такие психические феномены, как состояние и самочувствие.
В последующем, кроме активирующего влияния ретикулярной формации, при повреждении последней были описаны акинезы, сонливость, угнетенное состояние сознания. Наряду с влиянием ретикулярной формации на корковые функции было установлено наличие кортико-фугальных волокон, идущих из лобных отделов, поясной извилины, сенсомоторной коры и некоторых других ее областей к ретикулярной формации. С помощью электрофизиологических исследований было показано, что проведение импульсов повосходящим путям ретикулярной формации может быть заторможено или облегчено возбуждением вышеуказанных участков коры. Таким образом, корковая, в том числе и сознательная, деятельность может оказывать определенное влияние на активационно-регу-ляционные механизмы субкортикальных структур, обеспечивающих ее же (коры) функционирование.
Интенсивное изучение глубоких структур мозга позволило существенно расширить представления об анатомо-физиологическом субстрате эмоций. В экспериментах на «таламических» и «гипоталамических» (декортицированных) животных было показано, что моторное выражение эмоций может осуществляться и при отсутствии новой коры, однако, естественно, что при этом не возникает интеграции эмоций в целостный поведенческий акт и нет эмоциональных ощущений. В последующем изложенные факты были подтверждены клиническими наблюдениями над больными с поражением лобных долей. Эти наблюдения в значительной степени способствовали уточнению представлений о роли таламуса и гипоталамуса в эмоциональном поведении человека. Оказалось, что возникновение эмоциональных ощущений, выражение и регуляция их проявлений осуществляются за счет взаимодействия новой коры с гипоталамусом, лимби-ческой и ретикулярной системами, связи которых лежат в основе центрального нервного субстрата эмоций [Гельмгорн Э., Луфборроу Дж.]. Особую роль гипоталамуса в механизмах осуществления эмоционального поведения подчеркивал и P. Mac-Lean, сравнивая его с «водителем» (гипоталамус и обонятельный мозг) нервного «шасси» (спинной мозг и ствол), который выбирает возможные направления поведения в целом.
Сложившиеся в процессе эволюции тесные связи обонятельного мозга с двигательной, вегетативно-висцеральной, эндокринной и эмоциональной системами (J. Papez), с учетом общих принципов организации регуляторной деятельности коры головного мозга (через системы прямых и обратных связей), позволяет предполагать, что воздействие на одну из названных систем, например двигательную, неизбежно должно вызывать определенные изменения и в функционировании остальных. В свете последних достижений нейрофизиологии стало более понятным влияние мышечного расслабления на функции центральной и периферической нервной системы. Состояние релаксации вызывает снижение реактивности гипоталамуса, а следовательно, и эмоциональную возбудимость. И наоборот, снижение уровня центрального возбуждения в результате психотерапевтического (вербального, рационального) воздействия ведет к уменьшению нисходящих корковогипоталамо-соматических разрядов в системе обратной связи. Независимо от того, вызвано ли расслабление скелетной мускулатуры физиологическим, физиотерапевтическим, фармакологическим или психотерапевтическим воздействием, оно сопровождается снижением эмоциональной реактивности и ослаблением невротической симптоматики. Однако если физиологическое, физиотерапевтическое, фармакологическое и большинство видов психотерапевтического воздействия, оказывая существенное влияние на корковые и психические функции, реализуют свой терапевтический эффект при пассивном участии сознания, аутогенная тренировка предполагает активное вовлечение сознания в терапевтический процесс и осуществляется именно на основе активного сознания. Другими словами, аутогенная тренировка направлена на максимальное использование и стимуляцию резервных возможностей интегративной деятельности коры головного мозга на основе перестройки субординационных нервно-регуляторных корково-подкорково-спинальных механизмов.