ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 05.05.2024
Просмотров: 193
Скачиваний: 0
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.

F. evanescensфракцию фукоидана, содержащую приблизительно в 3.5 раза больше (1^3)-связанных остатков фукозы, чем (1^4)-связанных (Kusaykin et al., 2006). В обеих работах представлены данные моносахаридного состава фукоидана, содержащего помимо фукозы, минорные компоненты — ксилозу, маннозу и галактозу. При этом положение этих минорных компонентов в структуре фукоидана не было установлено. В нашей работе (Anastyuk et al., 2009) мы показали, что, используя тандемную масс-спектрометрию с ионизацией электрораспылением (ИЭР МС/МС), возможно установить структурные особенности не только сульфатированных фукоолигосахаридов, но и смешанных олигосахаридов, содержащих минорные моносахариды, что не всегда доступно методам ЯМР-спектроскопии.
Исследуемый нами образец фукоидана из F. evanescens(FeF2) был выделен по методу, описанному в работе (Kusaykin et al., 2006). В условиях сольволитического десульфатирования этого фукоидана помимо полимерной фракции был получен набор олигосахаридов, как в случае с фукоиданом из L. gurjanovae. Экстракцией этанолом из реакционной смеси была выделена доступная для масс-спектрометрического анализа олигосахаридная фракция Fe-dSlmf. По данным МАЛДИ МС положи-
78
тельных ионов, фракция Fe-dSlmf содержала набор интенсивных пиков, соответствующих ионам [Fuc +Na]+ (n=l-6) (см. рис.3.14, вставка). Спектр также содержал менее интенсивные сигналы, соответствующие смешанным олигосахаридам, в которых один или несколько остатков фукозы замещены остатком другого моносахарида, например, [Fuc1Pent1+Na]+ с mlz319.0 Да и [Fuc1Hex1+Na]+ с mlz349.1 Да, отстоящих от иона [Fuc2+Na]+ с mlz333.1 Да на -14 и +16 Да соответственно. Таким образом, несуль-фатированные олигосахариды, содержащие остатки пентозы и гексозы были найдены в МАЛДИ масс-спектре фракции Fe-dSlmf: [Fuc Pent+Na]+, п=1-6 и [Fuc Hex+Na]+, n=l-6.
220 240 260 280 300 320 340 360 380 400 420 440 460 480 500 520 540 560 580 600 620 640 660 680
Cojnts ys. Mass-to-Charge (rm/z)
Рис. 3.14. ИЭР масс-спектр сульфатированных олигосахаридов фракции Fe-AFl, полученной из Fe-dSlmf с помощью обращенно-фазовой ВЭЖХ. Вставка содержит МАЛДИ масс-спектр положительных ионов фракции Fe-dSlmf, полученной в условиях сольволитического десульфатирования
МАЛДИ масс-спектр, полученный в режиме регистрации отрицательных ионов, содержал набор интенсивных пиков ионов, соответствующих моносульфатированным фукозе и фукоолигомерам со степенью полимеризации 2—4 (данные не приведены). Это означало, что десульфатирование прошло не полностью и в смеси присутствовали сульфатированные компоненты. С целью обогащения смеси анионными компонентами фракция Fe-dSlmf была разделена с помощью обращенно-фазовой ВЭЖХ. Фракция 1 (Fe-AFl), не задержавшаяся на колонке, была проанализирована с помощью ИЭР МС в режиме регистрации отрицательных ионов. Высокая чувствительность тандемного ИЭР масс-спектрометра с времяпролетным анализатором позволила обнаружить
79
не только сульфатированные компоненты, но и фукоолигомеры, содержащие уроновую кислоту, для большей части из которых также были получены МС/МС спектры (см рис. 3.14). Согласно моносахаридному составу фракции Fe-AFl пентозе соответствует ксилоза (Xyl), гексо-зе — галактоза (Gal), уроновая кислота была идентифицирована как глюкуроновая (GlcA). Полная информация по составу данной фракции отображена в таблице 3.1.
Таблица 3.1 Значения m/zосновных и минорных (*) низкомолекулярных продуктов сольволитического десульфатирования (фракция Fe-AFl) фукой дана
из F. evanescensyполученные с помощью ИЭР МС
Наличие в фукоидане из F. evanescensостатков сульфатированных ксилозы (m/z229.006) и галактозы (m/z259.018), а также фрагментов, содержащих уроновую кислоту ([FucnGlcA-Na]", n=l-3), было показано нами впервые (Anastyuk et al, 2009).
Каждый ион, наблюдаемый в ИЭР масс-спектре (табл. 3.1), был подвергнут фрагментации с целью извлечения структурной информации. Стоит отметить, что отдельные сигналы ионов в масс-спектре могут соответствовать нескольким изомерам, имеющим одинаковую массу, но отличающимся положением сульфатных групп, типом связи, положением остатков различных моносахаридов и др. Однако, благодаря характеристичным фрагментам, нередко становится возможным установление структурных особенностей отдельных изомеров.
На рис. 3.15 представлена картина распада иона с m/z229.003, соот-
80

140 150 160 170 180 190 200 210 220 230 Counts vs. Mass-to-Charge (m/z)
ветствующего [XylS03Na-Na] . Интенсивные фрагменты с m/z138.970 и 168.979 соответствовали двойным разрывам связей внутри кольца. Первый ион 02Х свидетельствовал о 2-0-, а второй — 0,2А — о 4-О-сульфатировании остатка ксилозы. Следовательно, исходный полисахарид содержит 2-0- и 4-О-сульфатированные остатки ксилозы.
Тандемный масс-спектр иона фрагмента, содержащего кси-
400
600
800
1000
1200 m/z
Рис. 3.23. МАЛДИ масс-спектр отрицательных ионов олигосахаридов, полученных автогидролизом фукоидана из бурой водоросли F. evanescens
88
Структурные характеристики олигосахаридов были установлены с помощью МАЛДИ МС/МС. Основной компонент смеси — дисульфа-тированная фукобиоза имела преимущественно (1^4)-гликозидную связь между остатками фукозы (Anastyuk et al., 2012). Ее тандемный масс-спектр представлен на рисунке 3.24.
100J
гсбСН
0
Na03SO

°'2А2 gj
В]
о
Y,
°'2ХЛ
hso;
т- О)
I о
со
0,2Х,
В', аэ со
см
\
[M(2Na)-NaHS04-Na] I °<2Х',
т- СО h- CO СО СО
И»и,^1 ,.
jjkb.
+ "f4 '
М*
100 150 200 250 300 350 400 m/z
Рис. 3.24. Тандемный МАЛДИ масс-спектр иона [Fuc (SO Na)2-Na]" c m/z491.0
По характеру интенсивностей фрагментных ионов масс-спектр оказался более похож на тандемный масс-спектр дисульфатированной фукобио-
зы, выделенной из фукоидана А.
P^laOsSOv'

Рис. 3.25. Тандемный МАЛДИ масс-спектр иона [Fuc,(SO,Na),-Na]- с m/z593.0
nodosum, где авторами было отмечено существенное количество (1^4)-связанных остатков a-L-Fucp. (Daniel et al., 2007) При этом, из спектра не следовало, есть ли в смеси (1^3)-связанные остатки a-L-Fucp.
Этот вопрос был снят при анализе иона трисульфатированной фукобиозы с m/z593.0. (рис. 3.25). Благодаря наличию дополнительной сульфатной группы, мы смогли наблюдать Y-ион с m/z344.9, соответствующий отщеплению дважды сульфатированного остатка фукозы с восстанавливающего конца молекулы. Поскольку в фу-
89
коидане из F. evanescensсульфатирование при СЗ остатка ct-L-Fucp не было обнаружено (Bilan et al., 2002), был сделан вывод о присутствии некоторого количества (1^3)-связанных остатков ct-L-Fucp (рис. 3.24).
Остатки гексозы (D-Gal) были обнаружены в составе ди- и трисульфа-тированных дисахаридов типа FucHex. По результатам анализа тандемно-го масс-спектра (рис. 3.26), было предложено 3 варианта строения дисаха-рида: Gal-2-OS03Na-(l->3)-Gal-2-OS03Na, Gal-2,4-
Исследуемый нами образец фукоидана из F. evanescens(FeF2) был выделен по методу, описанному в работе (Kusaykin et al., 2006). В условиях сольволитического десульфатирования этого фукоидана помимо полимерной фракции был получен набор олигосахаридов, как в случае с фукоиданом из L. gurjanovae. Экстракцией этанолом из реакционной смеси была выделена доступная для масс-спектрометрического анализа олигосахаридная фракция Fe-dSlmf. По данным МАЛДИ МС положи-
78
тельных ионов, фракция Fe-dSlmf содержала набор интенсивных пиков, соответствующих ионам [Fuc +Na]+ (n=l-6) (см. рис.3.14, вставка). Спектр также содержал менее интенсивные сигналы, соответствующие смешанным олигосахаридам, в которых один или несколько остатков фукозы замещены остатком другого моносахарида, например, [Fuc1Pent1+Na]+ с mlz319.0 Да и [Fuc1Hex1+Na]+ с mlz349.1 Да, отстоящих от иона [Fuc2+Na]+ с mlz333.1 Да на -14 и +16 Да соответственно. Таким образом, несуль-фатированные олигосахариды, содержащие остатки пентозы и гексозы были найдены в МАЛДИ масс-спектре фракции Fe-dSlmf: [Fuc Pent+Na]+, п=1-6 и [Fuc Hex+Na]+, n=l-6.
| s 7,5 7- | | | | [Fuc2S03Na | -Na]'—| CO | [Fuc,+Na]l[Fuc'+Na]' [F"«V+-Na]* зззi 1 479'' S!5'' | | |||||
| 6,5- e 6- | [FucS03Na-Na] | | | | [Fuq +Na]' | | | | [Fuc5+Na]' 771 .Л | |||
| §5,5- | | 1 | | | | 187 2 | | | | | [Fuc6+Na]t | ||
| s 5-O | | Я ■ | | | | 1 1 1 | | | | | 517.5 | |
| £ 4- | | S | | | | | | 1 | „,,, 4 | L ..i | 1_____1^ | |
| | | | | |||||||||
| < 3- | | | | | | | | | ||||
| 2,5 2- | | | | | | | [Fuc3S03Na-Na]"—£ s 3 [Fuq,S03Na-Na] | |||||
| 1,5- 1 0,5 0- | О Oi CSI | _J | n о о" ш | 0> ■ -I \ | g S s , .ll.ll ll | CO * ОС о — Ч> — о ^_- « я ■Г "1 ■■!......A -f4 ' | i со _ — см сд ^-т г-" « щ 1 Й щ 1 1- L ™ ......i............1. | |||||
220 240 260 280 300 320 340 360 380 400 420 440 460 480 500 520 540 560 580 600 620 640 660 680
Cojnts ys. Mass-to-Charge (rm/z)
Рис. 3.14. ИЭР масс-спектр сульфатированных олигосахаридов фракции Fe-AFl, полученной из Fe-dSlmf с помощью обращенно-фазовой ВЭЖХ. Вставка содержит МАЛДИ масс-спектр положительных ионов фракции Fe-dSlmf, полученной в условиях сольволитического десульфатирования
МАЛДИ масс-спектр, полученный в режиме регистрации отрицательных ионов, содержал набор интенсивных пиков ионов, соответствующих моносульфатированным фукозе и фукоолигомерам со степенью полимеризации 2—4 (данные не приведены). Это означало, что десульфатирование прошло не полностью и в смеси присутствовали сульфатированные компоненты. С целью обогащения смеси анионными компонентами фракция Fe-dSlmf была разделена с помощью обращенно-фазовой ВЭЖХ. Фракция 1 (Fe-AFl), не задержавшаяся на колонке, была проанализирована с помощью ИЭР МС в режиме регистрации отрицательных ионов. Высокая чувствительность тандемного ИЭР масс-спектрометра с времяпролетным анализатором позволила обнаружить
79
не только сульфатированные компоненты, но и фукоолигомеры, содержащие уроновую кислоту, для большей части из которых также были получены МС/МС спектры (см рис. 3.14). Согласно моносахаридному составу фракции Fe-AFl пентозе соответствует ксилоза (Xyl), гексо-зе — галактоза (Gal), уроновая кислота была идентифицирована как глюкуроновая (GlcA). Полная информация по составу данной фракции отображена в таблице 3.1.
Таблица 3.1 Значения m/zосновных и минорных (*) низкомолекулярных продуктов сольволитического десульфатирования (фракция Fe-AFl) фукой дана
из F. evanescensyполученные с помощью ИЭР МС
| m/z | Состав | m/z | Состав | m/z | Состав |
| *229.000 | [XyljSOjNa-Na]" | 389.076 | [Fuc2S03Na-Na]" | 535.133 | [Fuc3S03Na-Na]" |
| 243.018 | [FuCjSOjNa-Na]" | *405.071 | [FuCjGaljSOjNa-Na]" | *551.128 | [FuCjGaljSOjNa-Na]" |
| *259.013 | [GaljSOjNa-Na]" | 421.060 | [Gal2S03Na-Na]" | *567.125 | [FuCjGaljSOjNa-Na]" |
| *339.093 | [Fuc^lcAJ- | 485.151 | [Fu^GlcAJ- | *631.209 | [FUC3GICAJ- |
| *375.060 | [FuCjXyljSOjNa-Na]" | *521.118 | [Fuc^CyljSOjNa-Na]- | *681.191 | [Fuc4S03Na-Na]" |
Наличие в фукоидане из F. evanescensостатков сульфатированных ксилозы (m/z229.006) и галактозы (m/z259.018), а также фрагментов, содержащих уроновую кислоту ([FucnGlcA-Na]", n=l-3), было показано нами впервые (Anastyuk et al, 2009).
Каждый ион, наблюдаемый в ИЭР масс-спектре (табл. 3.1), был подвергнут фрагментации с целью извлечения структурной информации. Стоит отметить, что отдельные сигналы ионов в масс-спектре могут соответствовать нескольким изомерам, имеющим одинаковую массу, но отличающимся положением сульфатных групп, типом связи, положением остатков различных моносахаридов и др. Однако, благодаря характеристичным фрагментам, нередко становится возможным установление структурных особенностей отдельных изомеров.
На рис. 3.15 представлена картина распада иона с m/z229.003, соот-
80
140 150 160 170 180 190 200 210 220 230 Counts vs. Mass-to-Charge (m/z)
ветствующего [XylS03Na-Na] . Интенсивные фрагменты с m/z138.970 и 168.979 соответствовали двойным разрывам связей внутри кольца. Первый ион 02Х свидетельствовал о 2-0-, а второй — 0,2А — о 4-О-сульфатировании остатка ксилозы. Следовательно, исходный полисахарид содержит 2-0- и 4-О-сульфатированные остатки ксилозы.
Тандемный масс-спектр иона фрагмента, содержащего кси-
Рис. 3.15. Тандемный ИЭР масс-спектр иона [XylS03Na-Na]- с m/zof 229.003 лозу [FuclXy^SC^Na-Na] с m/z375.059 (рис. 3.16), представлял более сложную картину.
Фрагментные ионы Z-типа с m/z210.993 и 225.006 были образованы вследствие разрыва гликозидных связей и являлись дегидратированными сульфатированными остатками ксилозы и фукозы соответственно. Остаток сульфатированной ксилозы, находящийся на восстанавливающем конце, давал менее интенсивные фрагментные ионы Yl-типа (значение m/zна спектре не отмечено), чем остаток сульфатированной фукозы с m/z243.014. Интенсивный фрагментный ион 0,2Х0 с m/z138.972 свидетельствовал о наличии сульфатной группы в основном при С2 остатков обоих моносахаридов, находящихся на восстанавли-
х1(Г
6 5,5
Л 5
8 4,5
I
g 4
О
н i 3
Ю 2,5 <
2
1,54
0.5Н

140 150 160 170 180 190 200 210 220 230 240 250 260 270 280 290 300 310 320 330 340 350 360 370 380 ITl/Z
Рис. 3.16. Тандемный ИЭР масс-спектр иона [XyljFuCjSC^Na-Na]" с m/z375.059
81
вающем конце. Однако, присутствие ионов 0,2А низкой интенсивности с m/z168.976 и 182.994 указывало на наличие сульфатных групп при С4 остатков ксилозы и фукозы, расположенных на невосстанавливающем конце молекулы. Наличие ионов 0,2Х с m/z271.008 и 285.032, а также 0,3Х1 с m/z301.012 говорило о 2-О-сульфатировании тех же остатков. Ионы °'2Х0 и °'2А2 с относительно высокой интенсивностью (последний ион возникает в случае присутствия в димере сульфатной группы на остатке моносахарида, находящегося на невосстанавливающем конце) свидетельствовали об (1^4)-связи между остатками фукозы и ксилозы. Попытка получить однозначные данные из МС/МС спектра иона трисахарида [Fuc^Xy^SC^Na-Na]" с mlz521.126 успехом не увенчалась.
На рис. 3.17 представлен МС/МС спектр иона [Fu^GlcA-Na]" с m/z339.093. Самый интенсивный фрагментный ион 0,2Х с m/z235.046 дает
хЮ
4,25-1
41 3,75^
3,5^
н 3,25
0 з
1 2,75^
Ч2,5
Р 2,25 I
ё 1,75
< 1,5
1,25^
U
0,75^
0,5^
0,25^
0^
[MNa-Na]"
-ОС 6 3 Y2 z2 он
'СН3 /\ц—^у3
x>hs
[MNa-H20-Na]"
180 200 220 240 260 280 300 320 340 360 380 400 420 440 460 480 500 520 540 560 580 600 620 640 m/z
Рис. 3.17. Тандемный ИЭР масс-спектр иона [FuCjGlcA-Na]" с m/z339.093
информацию о разрыве двух связей внутри кольца остатка фукозы, находящегося на невосстанавливающем конце. Вероятно, аномально высокая интенсивность этого фрагмента обусловлена присутствием в смеси GlcA-(l->2)-Fuc, когда фрагмент 0>2А имеет указанное m/z.
Фрагментные ионы, полученные в результате разрыва гликозидных связей и имеющие m/z193.036 и 175.025, были отнесены к ионам Z и Y , принадлежащим соответственно уроновой кислоте и ее дегидратированному остатку. Ион с m/z261.061 был отнесен к 2,5А2-типу, что характерно для уроновых кислот, находящихся на восстанавливающем конце (Zhang, et. al., 2006). Фрагментный ион 0,2А2 с m/z279.067, имеющий низкую интенсивность и, как упоминалось ранее, несущий информацию о типе свзязи, свидетельствовал о наличии в смеси преимущественно структур Fuc-
82
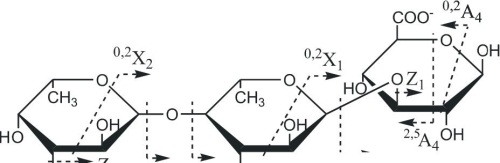
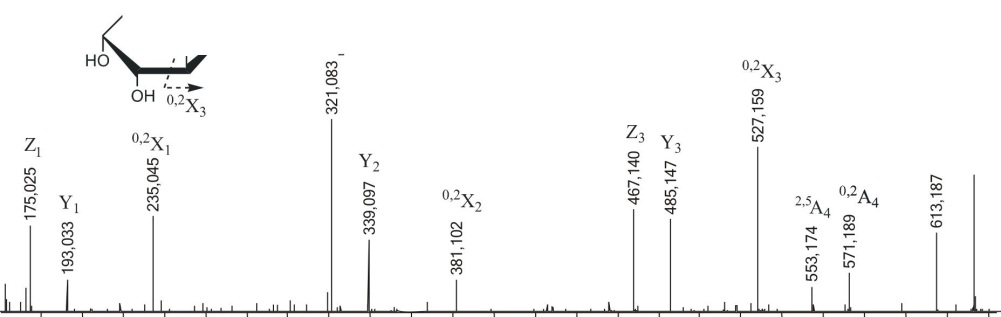
(l->3)-GlcA и GlcA-(1^2)-Fuc с небольшим количеством Fuc-(l-M)-GlcA. Фрагментационные картины ионов [Fuc2GlcA-Na] с m/z485.158 и [Fuc3GlcA-Na]" с m/z631.204 были аналогичными, поэтому нами дано описание МС/МС спектра более сложного иона (рис. 3.18).
хЮ2 4,25-1
4-3.75J 3.5 J
£3.25
l 3
^2,25 i8 1,75-1
< А
1.2sJ
1 0,7б|
o.sj
0,25 \
oJ-^
180 200 220 240 260 260 300 320 340 360 380 400 420 440 460 480 500 520 540 560 580 600 620 640
Courts vs. Mass-to-Charge (m/z)
Рис. 3.18. Тандемный ИЭР масс-спектр иона [Fuc3GlcA-Na]" с mlz631.204
Спектр содержал интенсивные ионы, соответствующие разрывам гликозидных связей. Ионы Z3 и Y3 с m/z467.140 и 485.147 соответствовали отщеплению с невосстанавливающего конца фукозы и ее дегидратированной формы; ионы Z2 и Y2 с m/z321.083 и 339.097 — фукобиозы и ее дегидратированной формы и т.д. Спектр также содержал фрагментные ионы 0,2А4 и 0,3А4 с m/z553.174 и 571.189, характерные для фрагментации уроновой кислоты, находящейся на восстанавливающем конце (см. описание МС/МС спектра иона [Fu^GlcA-Na]") и не содержал ионов °'3А или °'3Х, которые свидетельствуют о наличии разветвлений. Анализ спектра позволил сделать вывод о линейном строении данного фрагмента, имеющего уроновую кислоту на восстанавливающем конце. Отсутствие сульфатных групп, влияющих, по-видимому, на интенсивность сигналов 0,2Х, а также определенное положение уроновой кислоты позволило полуколичественно оценить 0>2Х-фрагменты, полученные из остатка фукотриозы, сравнивая их интенсивности. Фрагментный ион 0>2Х3 с mlz527.159, соответствующий двойному разрыву кольца фукозы на восстанавливающем конце, имел наибольшую интенсивность из-за наличия незамещенного протона в гидроксиле при СЗ, участвующего в механизме образования фрагментных ионов данного типа, как уже было сказано выше. Фрагментный ион 0>2Х2 с m/z381.102, принадлежащий второму остатку фукозы с восстанавливающего конца, имел значительно
83
меньшую интенсивность, что указывало на (1^3)-связь между остатками фукозы. Интенсивность следующего фрагмента 0,2Х1 с m/z235.045, образующегося из третьего по счету с невосстанавливающего конца остатка фукозы, была примерно в 3 раза выше, указывая на преобладание (1^4)-связи между третьим и соседним с ним остатком фукозы. Низкая интенсивность фрагментного иона 0,2А4 уроновой кислоты с m/z571.204 также свидетельствовала о преимуществе (1^3)-связи между остатками фукозы в этой позиции.
На основании МС/МС анализа можно представить наиболее вероятную структуруданногоолигомеракакРис-(1^3)-Рис-(1^4)-Бис-(1^3)-С1сА. Подобный тандемный масс-спектр иона [Fuc2GlcA-Na]" с m/z485.151 (данные не приведены) свидетельствовал о преобладании структуры Fuc-(l->4)-Fuc-(l->3)-GlcA. Это вполне согласуется со структурой фракции фукоидана из Е evanescens, опубликованной в работе (Bilan et al., 2002), которая представляет собой линейную цепь, построенную из сульфатированных остатков фукозы с чередующимися (1->3)- и (1->4)-связями. Фрагменты, представляющие фукоолигосахариды, имеющие глюкуроновую кислоту в своем составе, обнаруженные нами в фукоидане из F. evanescens, построены подобным образом.
Анализ фрагментационной картины интенсивного иона [FucS03Na-Na]
с m/z243.016 (данные не приведены) дал результаты, согласующиеся с полученными ранее для A. nodosum(Daniel et al., 2007). Интенсивность иона 0>2А с m/z182.996 (фрагмент 4-О-сульфатированной фукозы) была примерно в 2 раза ниже, чем интенсивность иона 0,2Х с m/z138.971 (фрагмент 2-О-сульфатированной фукозы). Полученные результаты согласуются с литературными данными (Bilan et al., 2002, Kusaykin et al., 2006), из которых следует, что остатки фукозы в в фукоидане из Е evanescensсульфатированы в основном при С2.
Фрагментационные картины наиболее интенсивного иона [Fuc2S03Na-Na]
с m/z389.082 и иона [Fuc3S03Na-Na]
с m/z535.131 были похожи, поэтому мы привели анализ МС/МС спектра последнего иона (рис. 3.19). Наиболее интенсивными ионами МС/МС спектра были сульфатированная фукоза (m/z243.017) и ее дегидратированная форма (m/z225.007), образовавшиеся в результате разрыва гликозидных связей.
В спектре были найдены менее интенсивные фрагментные ионы с m/z389.074 и 371.065, соответствующие сульфатированной фукобиозе и ее дегидратированной форме. Кроме того, имелись сигналы ионов °'2Х0 и 0,2Х , получающиеся в результате двойного разрыва моносахаридного кольца с m/z138.971 и с m/z285.029, которые также давали информацию о сульфатировании гидроксилов при С2 остатков фукозы на восстанав-
84
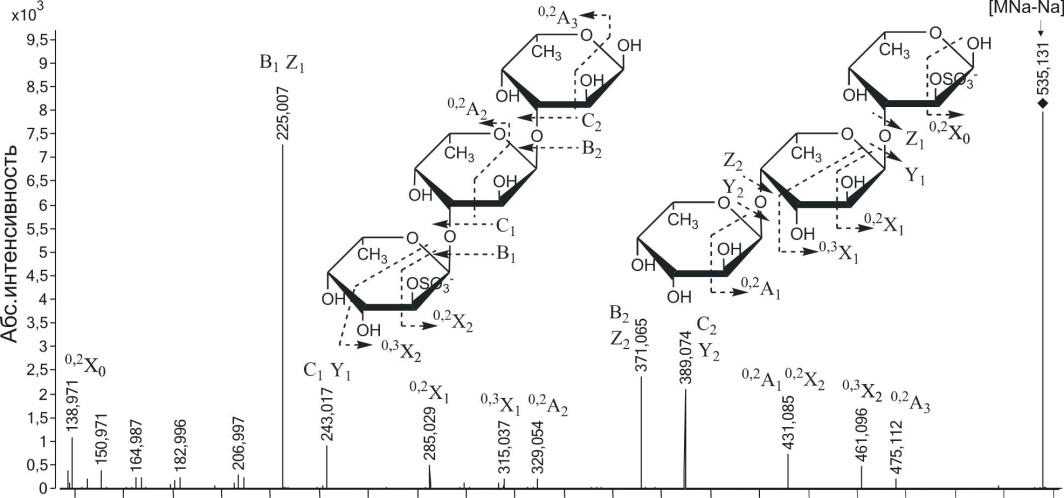
Рис. 3.19. Тандемный ИЭР масс-спектр иона [Fuc3S03Na-Na]- с m/z535.131
ливающем конце и соседнего с ним. Фрагментный ион 0,2Х2 с m/z431.085 возникал из сульфатированного при С2 остатка фукозы, находящегося на невосстанавливающем конце. Присутствие фрагментного иона малой интенсивности с m/z182.996 свидетельствовало о 4-О-сульфатировании невосстанавливающего остатка фукозы. Фрагментные ионы, несущие информацию о 4-О-сульфатировании других остатков фукозы в оли-гомере, обнаружены не были. Ион °'2А2 низкой интенсивности с m/z329.054 (по сравнению с ионом 0>2Х2 с m/z431.085, полученным из невосстанавливающего остатка со свободным гидроксилом при СЗ) давал информацию о вероятности (1^3)-связи между остатками фукозы на невосстанавливающем конце (рис. 3.19, слева). Однако, МС/МС спектр содержал ионы 0,3Х1 с m/z315.037 и 0,2Х1 с m/z285.029, указывающие на наличие (1^4)-связи между этими же остатками (рис. 3.19, справа). Ион очень низкой интенсивности 0,2А3 с m/z475.112 и вышеупомянутый 0,2Х0 свидетельствовали о вероятности (1^3)-связи между остатками фукозы, находящимися на восстанавливающем конце. Таким образом, методом МС/МС обнаружены фрагменты фукоидана, построенные из сульфатированных остатков фукозы с чередующимися (1-»3)- и (1->4)-связями, что также находится в соответствии с результатами предыдущих исследований (Kusaykin et al., 2006). Анализ более протяженных сульфатированных фукоолигосахаридов не дал столь четких результатов из-за большого количества шума в спектре и сложности в интерпретации.
Использование тандемной масс-спектрометрии позволило установить структурное значение в фукоидане из F. evanescensеще одного минорного компонента - галактозы. МС/МС спектр иона с m/z259.018, соответствующего [GalS03Na-Na]", давал информацию о С2 и С4/С6 сульфатировании
85
хЮ 3.4-1
3,2^
3 I 2,8^
2,6^
^2,4
8 2,2
I
1 2
ff 1.6
ii,4
ё1,2 < 1
0,8^ 0,6^ 0/N
0,2^
oJ
CH2OH ' Al CH2OH
-о//"*" Л-—ov/

Hid ,.il 1.1
MX, 0,2Y
CH2OH ,.*J CH2OH _A^
-О'' V, 7 J--------О /
■\ .J- r2/ К ОН
°>3X,
[MNa-Na]"
130 140 150 160 170 180 190 200 210 220 230 240 250 260 270 280 290 300 310 320 330 340 350 360 370 380 390 400 410 420 430 m/Z
Рис. 3.20. Тандемный ИЭР масс-спектр иона [Gal2S03Na-Na]" с m/z421.060
остатков галактозы. МС/МС спектр иона [Gal2SO Na-Na] с m/z421.060 (рис. 3.20) содержал интенсивный фрагментный ион с m/z241.000 и менее интенсивный с m/z259.012. Образование этих ионов связано с разрывом гликозидных связей и отщеплением дегидратированного остатка сульфа-тированной галактозы и сульфатированной галактозы соответственно.
Ион 0,2Х0 с m/z138.970 указывал на сульфатирование остатка галактозы на восстанавливающем конце при С2. Фрагментные ионы 0,2Х с m/z301.022 и 0,3Х1 с m/z331.032 свидетельствовали о 2-О-сульфатировании остатка галактозы, находящегося на невосстанавливающем конце. Фрагменты, характерные для сульфатирования остатка галактозы при С4, отсутствовали. Таким образом, МС/МС-анализ позволил обнаружить и установить структуру галактозосодержащих фрагментов фукоидана: Gal2S03--(l-H)-Gal и Gal-(l->4)-Gal2S03.
Галактоза входила также в состав смешанного дисахарида [Fu^Ga^SO^a-Na] с m/z405.070, присутствующего в МС-спектре. Как и в случае с предыдущими сульфатированными дисахаридами, этот ион содержал несколько изомеров, различающихся положением в них моно-сахаридных остатков и сульфатных групп. МС/МС спектр (рис. 3.21) содержал интенсивные ионы с m/z225.007 и 241.003, образование которых связано с разрывом гликозидных связей и отрывам сульфатированных остатков дегидратированной фукозы и галактозы. Фрагментные ионы 0,2Х-типа с m/z285.029 и 301.021 указывали на 2-О-сульфатирование невосстанавливающих остатков фукозы и галактозы.
Фрагментный ион оаА с m/z198.992 (на рисунке не отмечен) указывал на 4-О-сульфатирование невосстанавливающего остатка галактозы,
86
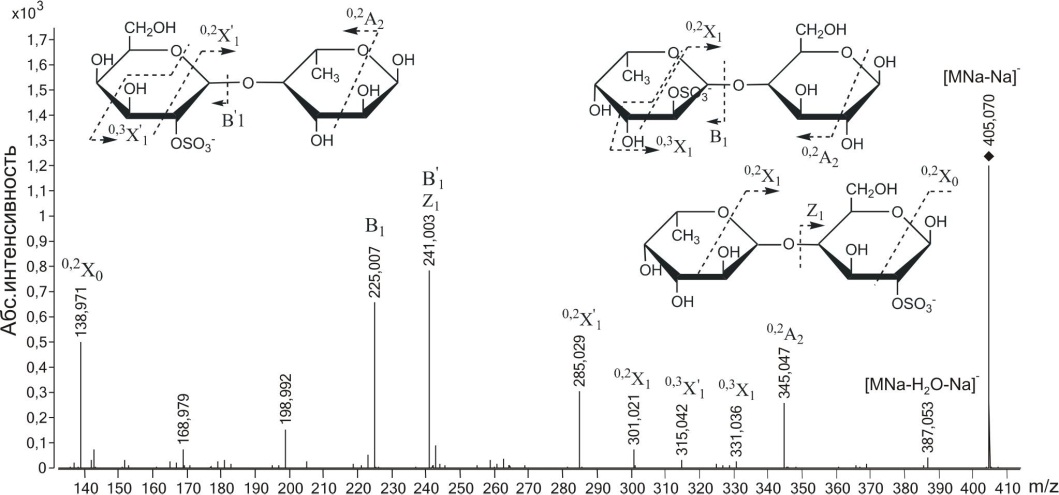
Рис. 3.21. Тандемный ИЭР масс-спектр иона [FuCjGaljSC^Na-Na]- с m/z405.070
следов 4-О-сульфатирования остатков фукозы не обнаружено. Присутствие ионов 0,2А2 с m/z345.047 и 0,2Х0, указывает на наличие (1^4)-связи, однако наличие (1-*3) -связи не может быть исключено, т.к. в таком случае, как уже было неоднократно отмечено, не образуются фрагментные ионы 0>2Х и 0>2А. Наличие иона 0>3Х0 с m/z168.979 также подтверждает присутствие структурного варианта Fuc-(1^4)-Gal-2-S03- в исследуемой смеси, т.к. для остатков сульфатированной галактозы вероятность фрагментации по типу 0,3Х была выше, чем для остатков фукозы (см. описание МС/МС спектра сульфатированной галактозы). Таким образом, масс-спектрометрический анализ показал наличие в смеси ионов Gal-(1^4)-Fuc-2-S03-, Gal-2-S03"-(l-H)-Fuc и незначительное количество Fuc-(1^4)-Gal-2-S03. Остаток галактозы на невосстанавливающем конце может быть сульфатирован при С4.
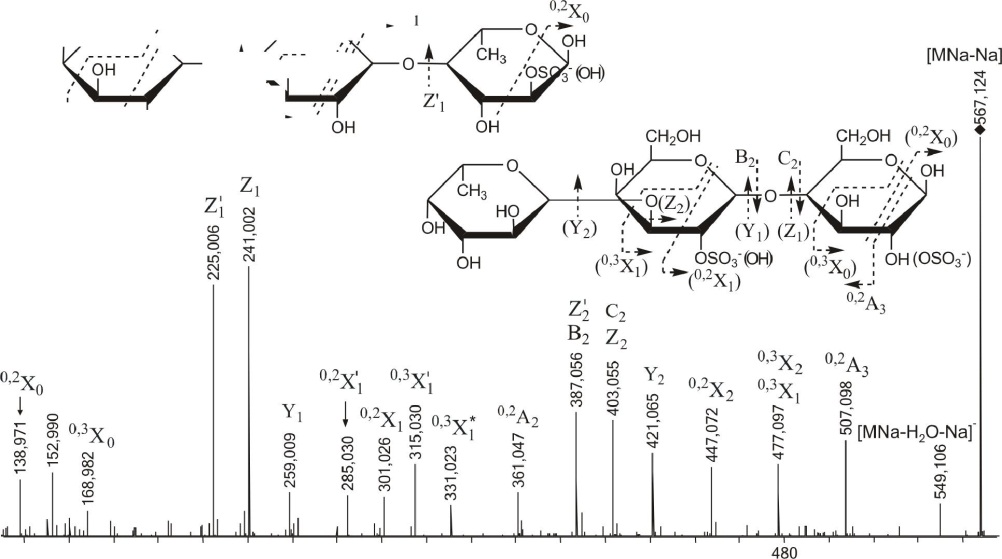
Картина масс-спектрометрической фрагментации иона [Fuc1Gal2S03Na--Na]
с m/z567.124 (см. рис. 3.22) была наиболее сложной для интерпретации. Тем не менее, спектр был интерпретирован аналогично предыдущим.
В результате был сделан вывод о превалирующих структурах исследуемого иона: Gal-(l-M)-Gal-(l-»3)-Fuc и Fuc-(l->3)-Gal-(l->4)-Gal, сульфатированные в основном при С2 остатков фукозы и галактозы, иногда — при С4 остатка галактозы.
Исследование фукоидана из F. evanescensбыло продолжено с использованием более мягких условий деполимеризации для получения мульти-сульфатированных фрагментов. Результаты исследования мультисульфа-тированных фрагментов фукоидана из F. evanescens, представлены в работе (Anastyuk et al, 2012). Необходимо отметить, что фукоидан был выделен нами в более «мягких» условиях, чем в работе (Kusaykin et al, 2006). С по-
87
хЮ 4 3,8 3,6 3,4 3,2
л 3
52,8
О 2,6
| 2,4
О 2,2
5 2 x 1,8
6 1.6
51,4
1.2Ч
(°'2A2)
сн2он о,2Х сн2он"*'! 0,2X он/— °-ч(^2 Л—Олл* -°-^С'он""77
Z'2L
0,3^" OH(OS03") 0,3X
140 160 180 200 220 240 260 280 300 320 340 360 380 400 420 440 460
500 520 540 560 m/z
Рис. 3.22. Тандемный ИЭР масс-спектр иона [FuCjGa^SC^Na-Na]" с m/z567.124. Структурные варианты с иным расположением сульфатных групп и соответствующие
фрагменты заключены в скобки
мощью автогидролиза (5 мг/мл) мы получили смесь олигосахаридов, состав которой был определен с помощью МАЛДИ МС. Смесь состояла из моно-сульфатированной фукозы и набора фукоолигосахаридов с четной степенью полимеризации (2—6), с числом сульфатных групп до 5. Олигосахариды, содержащие минорные моносахариды (в данном случае — галактозу, исходя из моносахаридного состава смеси), также были обнаружены (Рис. 3.23).
100
50
Lu.
hJ-j U
Рис. 3.15. Тандемный ИЭР масс-спектр иона [XylS03Na-Na]- с m/zof 229.003 лозу [FuclXy^SC^Na-Na] с m/z375.059 (рис. 3.16), представлял более сложную картину.
Фрагментные ионы Z-типа с m/z210.993 и 225.006 были образованы вследствие разрыва гликозидных связей и являлись дегидратированными сульфатированными остатками ксилозы и фукозы соответственно. Остаток сульфатированной ксилозы, находящийся на восстанавливающем конце, давал менее интенсивные фрагментные ионы Yl-типа (значение m/zна спектре не отмечено), чем остаток сульфатированной фукозы с m/z243.014. Интенсивный фрагментный ион 0,2Х0 с m/z138.972 свидетельствовал о наличии сульфатной группы в основном при С2 остатков обоих моносахаридов, находящихся на восстанавли-
х1(Г
6 5,5
Л 5
8 4,5
I
g 4
О
н i 3
Ю 2,5 <
2
1,54
0.5Н
140 150 160 170 180 190 200 210 220 230 240 250 260 270 280 290 300 310 320 330 340 350 360 370 380 ITl/Z
Рис. 3.16. Тандемный ИЭР масс-спектр иона [XyljFuCjSC^Na-Na]" с m/z375.059
81
вающем конце. Однако, присутствие ионов 0,2А низкой интенсивности с m/z168.976 и 182.994 указывало на наличие сульфатных групп при С4 остатков ксилозы и фукозы, расположенных на невосстанавливающем конце молекулы. Наличие ионов 0,2Х с m/z271.008 и 285.032, а также 0,3Х1 с m/z301.012 говорило о 2-О-сульфатировании тех же остатков. Ионы °'2Х0 и °'2А2 с относительно высокой интенсивностью (последний ион возникает в случае присутствия в димере сульфатной группы на остатке моносахарида, находящегося на невосстанавливающем конце) свидетельствовали об (1^4)-связи между остатками фукозы и ксилозы. Попытка получить однозначные данные из МС/МС спектра иона трисахарида [Fuc^Xy^SC^Na-Na]" с mlz521.126 успехом не увенчалась.
На рис. 3.17 представлен МС/МС спектр иона [Fu^GlcA-Na]" с m/z339.093. Самый интенсивный фрагментный ион 0,2Х с m/z235.046 дает
хЮ
4,25-1
41 3,75^
3,5^
н 3,25
0 з
1 2,75^
Ч2,5
Р 2,25 I
ё 1,75
< 1,5
1,25^
U
0,75^
0,5^
0,25^
0^
[MNa-Na]"
-ОС 6 3 Y2 z2 он
'СН3 /\ц—^у3
x>hs
[MNa-H20-Na]"
180 200 220 240 260 280 300 320 340 360 380 400 420 440 460 480 500 520 540 560 580 600 620 640 m/z
Рис. 3.17. Тандемный ИЭР масс-спектр иона [FuCjGlcA-Na]" с m/z339.093
информацию о разрыве двух связей внутри кольца остатка фукозы, находящегося на невосстанавливающем конце. Вероятно, аномально высокая интенсивность этого фрагмента обусловлена присутствием в смеси GlcA-(l->2)-Fuc, когда фрагмент 0>2А имеет указанное m/z.
Фрагментные ионы, полученные в результате разрыва гликозидных связей и имеющие m/z193.036 и 175.025, были отнесены к ионам Z и Y , принадлежащим соответственно уроновой кислоте и ее дегидратированному остатку. Ион с m/z261.061 был отнесен к 2,5А2-типу, что характерно для уроновых кислот, находящихся на восстанавливающем конце (Zhang, et. al., 2006). Фрагментный ион 0,2А2 с m/z279.067, имеющий низкую интенсивность и, как упоминалось ранее, несущий информацию о типе свзязи, свидетельствовал о наличии в смеси преимущественно структур Fuc-
82
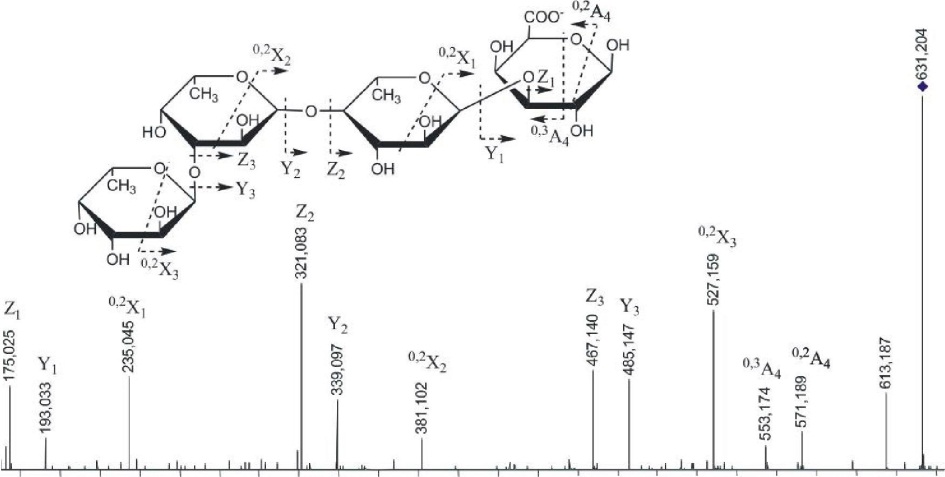
(l->3)-GlcA и GlcA-(1^2)-Fuc с небольшим количеством Fuc-(l-M)-GlcA. Фрагментационные картины ионов [Fuc2GlcA-Na] с m/z485.158 и [Fuc3GlcA-Na]" с m/z631.204 были аналогичными, поэтому нами дано описание МС/МС спектра более сложного иона (рис. 3.18).
хЮ2 4,25-1
4-3.75J 3.5 J
£3.25
l 3
^2,25 i8 1,75-1
< А
1.2sJ
1 0,7б|
o.sj
0,25 \
oJ-^
180 200 220 240 260 260 300 320 340 360 380 400 420 440 460 480 500 520 540 560 580 600 620 640
Courts vs. Mass-to-Charge (m/z)
Рис. 3.18. Тандемный ИЭР масс-спектр иона [Fuc3GlcA-Na]" с mlz631.204
Спектр содержал интенсивные ионы, соответствующие разрывам гликозидных связей. Ионы Z3 и Y3 с m/z467.140 и 485.147 соответствовали отщеплению с невосстанавливающего конца фукозы и ее дегидратированной формы; ионы Z2 и Y2 с m/z321.083 и 339.097 — фукобиозы и ее дегидратированной формы и т.д. Спектр также содержал фрагментные ионы 0,2А4 и 0,3А4 с m/z553.174 и 571.189, характерные для фрагментации уроновой кислоты, находящейся на восстанавливающем конце (см. описание МС/МС спектра иона [Fu^GlcA-Na]") и не содержал ионов °'3А или °'3Х, которые свидетельствуют о наличии разветвлений. Анализ спектра позволил сделать вывод о линейном строении данного фрагмента, имеющего уроновую кислоту на восстанавливающем конце. Отсутствие сульфатных групп, влияющих, по-видимому, на интенсивность сигналов 0,2Х, а также определенное положение уроновой кислоты позволило полуколичественно оценить 0>2Х-фрагменты, полученные из остатка фукотриозы, сравнивая их интенсивности. Фрагментный ион 0>2Х3 с mlz527.159, соответствующий двойному разрыву кольца фукозы на восстанавливающем конце, имел наибольшую интенсивность из-за наличия незамещенного протона в гидроксиле при СЗ, участвующего в механизме образования фрагментных ионов данного типа, как уже было сказано выше. Фрагментный ион 0>2Х2 с m/z381.102, принадлежащий второму остатку фукозы с восстанавливающего конца, имел значительно
83
меньшую интенсивность, что указывало на (1^3)-связь между остатками фукозы. Интенсивность следующего фрагмента 0,2Х1 с m/z235.045, образующегося из третьего по счету с невосстанавливающего конца остатка фукозы, была примерно в 3 раза выше, указывая на преобладание (1^4)-связи между третьим и соседним с ним остатком фукозы. Низкая интенсивность фрагментного иона 0,2А4 уроновой кислоты с m/z571.204 также свидетельствовала о преимуществе (1^3)-связи между остатками фукозы в этой позиции.
На основании МС/МС анализа можно представить наиболее вероятную структуруданногоолигомеракакРис-(1^3)-Рис-(1^4)-Бис-(1^3)-С1сА. Подобный тандемный масс-спектр иона [Fuc2GlcA-Na]" с m/z485.151 (данные не приведены) свидетельствовал о преобладании структуры Fuc-(l->4)-Fuc-(l->3)-GlcA. Это вполне согласуется со структурой фракции фукоидана из Е evanescens, опубликованной в работе (Bilan et al., 2002), которая представляет собой линейную цепь, построенную из сульфатированных остатков фукозы с чередующимися (1->3)- и (1->4)-связями. Фрагменты, представляющие фукоолигосахариды, имеющие глюкуроновую кислоту в своем составе, обнаруженные нами в фукоидане из F. evanescens, построены подобным образом.
Анализ фрагментационной картины интенсивного иона [FucS03Na-Na]
с m/z243.016 (данные не приведены) дал результаты, согласующиеся с полученными ранее для A. nodosum(Daniel et al., 2007). Интенсивность иона 0>2А с m/z182.996 (фрагмент 4-О-сульфатированной фукозы) была примерно в 2 раза ниже, чем интенсивность иона 0,2Х с m/z138.971 (фрагмент 2-О-сульфатированной фукозы). Полученные результаты согласуются с литературными данными (Bilan et al., 2002, Kusaykin et al., 2006), из которых следует, что остатки фукозы в в фукоидане из Е evanescensсульфатированы в основном при С2.
Фрагментационные картины наиболее интенсивного иона [Fuc2S03Na-Na]
с m/z389.082 и иона [Fuc3S03Na-Na]
с m/z535.131 были похожи, поэтому мы привели анализ МС/МС спектра последнего иона (рис. 3.19). Наиболее интенсивными ионами МС/МС спектра были сульфатированная фукоза (m/z243.017) и ее дегидратированная форма (m/z225.007), образовавшиеся в результате разрыва гликозидных связей.
Рис. 3.15. Тандемный ИЭР масс-спектр иона [XylS03Na-Na]- с m/zof 229.003 лозу [FuclXy^SC^Na-Na] с m/z375.059 (рис. 3.16), представлял более сложную картину.
Фрагментные ионы Z-типа с m/z210.993 и 225.006 были образованы вследствие разрыва гликозидных связей и являлись дегидратированными сульфатированными остатками ксилозы и фукозы соответственно. Остаток сульфатированной ксилозы, находящийся на восстанавливающем конце, давал менее интенсивные фрагментные ионы Yl-типа (значение m/zна спектре не отмечено), чем остаток сульфатированной фукозы с m/z243.014. Интенсивный фрагментный ион 0,2Х0 с m/z138.972 свидетельствовал о наличии сульфатной группы в основном при С2 остатков обоих моносахаридов, находящихся на восстанавли-
х1(Г
6 5,5
Л 5
8 4,5
I
g 4
О
н i 3
Ю 2,5 <
2
1,54
0.5Н
140 150 160 170 180 190 200 210 220 230 240 250 260 270 280 290 300 310 320 330 340 350 360 370 380 ITl/Z
Рис. 3.16. Тандемный ИЭР масс-спектр иона [XyljFuCjSC^Na-Na]" с m/z375.059
81
вающем конце. Однако, присутствие ионов 0,2А низкой интенсивности с m/z168.976 и 182.994 указывало на наличие сульфатных групп при С4 остатков ксилозы и фукозы, расположенных на невосстанавливающем конце молекулы. Наличие ионов 0,2Х с m/z271.008 и 285.032, а также 0,3Х1 с m/z301.012 говорило о 2-О-сульфатировании тех же остатков. Ионы °'2Х0 и °'2А2 с относительно высокой интенсивностью (последний ион возникает в случае присутствия в димере сульфатной группы на остатке моносахарида, находящегося на невосстанавливающем конце) свидетельствовали об (1^4)-связи между остатками фукозы и ксилозы. Попытка получить однозначные данные из МС/МС спектра иона трисахарида [Fuc^Xy^SC^Na-Na]" с mlz521.126 успехом не увенчалась.
На рис. 3.17 представлен МС/МС спектр иона [Fu^GlcA-Na]" с m/z339.093. Самый интенсивный фрагментный ион 0,2Х с m/z235.046 дает
хЮ
4,25-1
41 3,75^
3,5^
н 3,25
0 з
1 2,75^
Ч2,5
Р 2,25 I
ё 1,75
< 1,5
1,25^
U
0,75^
0,5^
0,25^
0^
[MNa-Na]"
-ОС 6 3 Y2 z2 он
'СН3 /\ц—^у3
x>hs
[MNa-H20-Na]"
180 200 220 240 260 280 300 320 340 360 380 400 420 440 460 480 500 520 540 560 580 600 620 640 m/z
Рис. 3.17. Тандемный ИЭР масс-спектр иона [FuCjGlcA-Na]" с m/z339.093
информацию о разрыве двух связей внутри кольца остатка фукозы, находящегося на невосстанавливающем конце. Вероятно, аномально высокая интенсивность этого фрагмента обусловлена присутствием в смеси GlcA-(l->2)-Fuc, когда фрагмент 0>2А имеет указанное m/z.
Фрагментные ионы, полученные в результате разрыва гликозидных связей и имеющие m/z193.036 и 175.025, были отнесены к ионам Z и Y , принадлежащим соответственно уроновой кислоте и ее дегидратированному остатку. Ион с m/z261.061 был отнесен к 2,5А2-типу, что характерно для уроновых кислот, находящихся на восстанавливающем конце (Zhang, et. al., 2006). Фрагментный ион 0,2А2 с m/z279.067, имеющий низкую интенсивность и, как упоминалось ранее, несущий информацию о типе свзязи, свидетельствовал о наличии в смеси преимущественно структур Fuc-
82
(l->3)-GlcA и GlcA-(1^2)-Fuc с небольшим количеством Fuc-(l-M)-GlcA. Фрагментационные картины ионов [Fuc2GlcA-Na] с m/z485.158 и [Fuc3GlcA-Na]" с m/z631.204 были аналогичными, поэтому нами дано описание МС/МС спектра более сложного иона (рис. 3.18).
хЮ2 4,25-1
4-3.75J 3.5 J
£3.25
l 3
^2,25 i8 1,75-1
< А
1.2sJ
1 0,7б|
o.sj
0,25 \
oJ-^
180 200 220 240 260 260 300 320 340 360 380 400 420 440 460 480 500 520 540 560 580 600 620 640
Courts vs. Mass-to-Charge (m/z)
Рис. 3.18. Тандемный ИЭР масс-спектр иона [Fuc3GlcA-Na]" с mlz631.204
Спектр содержал интенсивные ионы, соответствующие разрывам гликозидных связей. Ионы Z3 и Y3 с m/z467.140 и 485.147 соответствовали отщеплению с невосстанавливающего конца фукозы и ее дегидратированной формы; ионы Z2 и Y2 с m/z321.083 и 339.097 — фукобиозы и ее дегидратированной формы и т.д. Спектр также содержал фрагментные ионы 0,2А4 и 0,3А4 с m/z553.174 и 571.189, характерные для фрагментации уроновой кислоты, находящейся на восстанавливающем конце (см. описание МС/МС спектра иона [Fu^GlcA-Na]") и не содержал ионов °'3А или °'3Х, которые свидетельствуют о наличии разветвлений. Анализ спектра позволил сделать вывод о линейном строении данного фрагмента, имеющего уроновую кислоту на восстанавливающем конце. Отсутствие сульфатных групп, влияющих, по-видимому, на интенсивность сигналов 0,2Х, а также определенное положение уроновой кислоты позволило полуколичественно оценить 0>2Х-фрагменты, полученные из остатка фукотриозы, сравнивая их интенсивности. Фрагментный ион 0>2Х3 с mlz527.159, соответствующий двойному разрыву кольца фукозы на восстанавливающем конце, имел наибольшую интенсивность из-за наличия незамещенного протона в гидроксиле при СЗ, участвующего в механизме образования фрагментных ионов данного типа, как уже было сказано выше. Фрагментный ион 0>2Х2 с m/z381.102, принадлежащий второму остатку фукозы с восстанавливающего конца, имел значительно
83
меньшую интенсивность, что указывало на (1^3)-связь между остатками фукозы. Интенсивность следующего фрагмента 0,2Х1 с m/z235.045, образующегося из третьего по счету с невосстанавливающего конца остатка фукозы, была примерно в 3 раза выше, указывая на преобладание (1^4)-связи между третьим и соседним с ним остатком фукозы. Низкая интенсивность фрагментного иона 0,2А4 уроновой кислоты с m/z571.204 также свидетельствовала о преимуществе (1^3)-связи между остатками фукозы в этой позиции.
На основании МС/МС анализа можно представить наиболее вероятную структуруданногоолигомеракакРис-(1^3)-Рис-(1^4)-Бис-(1^3)-С1сА. Подобный тандемный масс-спектр иона [Fuc2GlcA-Na]" с m/z485.151 (данные не приведены) свидетельствовал о преобладании структуры Fuc-(l->4)-Fuc-(l->3)-GlcA. Это вполне согласуется со структурой фракции фукоидана из Е evanescens, опубликованной в работе (Bilan et al., 2002), которая представляет собой линейную цепь, построенную из сульфатированных остатков фукозы с чередующимися (1->3)- и (1->4)-связями. Фрагменты, представляющие фукоолигосахариды, имеющие глюкуроновую кислоту в своем составе, обнаруженные нами в фукоидане из F. evanescens, построены подобным образом.
Анализ фрагментационной картины интенсивного иона [FucS03Na-Na]
с m/z243.016 (данные не приведены) дал результаты, согласующиеся с полученными ранее для A. nodosum(Daniel et al., 2007). Интенсивность иона 0>2А с m/z182.996 (фрагмент 4-О-сульфатированной фукозы) была примерно в 2 раза ниже, чем интенсивность иона 0,2Х с m/z138.971 (фрагмент 2-О-сульфатированной фукозы). Полученные результаты согласуются с литературными данными (Bilan et al., 2002, Kusaykin et al., 2006), из которых следует, что остатки фукозы в в фукоидане из Е evanescensсульфатированы в основном при С2.Фрагментационные картины наиболее интенсивного иона [Fuc2S03Na-Na]
В спектре были найдены менее интенсивные фрагментные ионы с m/z389.074 и 371.065, соответствующие сульфатированной фукобиозе и ее дегидратированной форме. Кроме того, имелись сигналы ионов °'2Х0 и 0,2Х , получающиеся в результате двойного разрыва моносахаридного кольца с m/z138.971 и с m/z285.029, которые также давали информацию о сульфатировании гидроксилов при С2 остатков фукозы на восстанав-
84
Рис. 3.19. Тандемный ИЭР масс-спектр иона [Fuc3S03Na-Na]- с m/z535.131
ливающем конце и соседнего с ним. Фрагментный ион 0,2Х2 с m/z431.085 возникал из сульфатированного при С2 остатка фукозы, находящегося на невосстанавливающем конце. Присутствие фрагментного иона малой интенсивности с m/z182.996 свидетельствовало о 4-О-сульфатировании невосстанавливающего остатка фукозы. Фрагментные ионы, несущие информацию о 4-О-сульфатировании других остатков фукозы в оли-гомере, обнаружены не были. Ион °'2А2 низкой интенсивности с m/z329.054 (по сравнению с ионом 0>2Х2 с m/z431.085, полученным из невосстанавливающего остатка со свободным гидроксилом при СЗ) давал информацию о вероятности (1^3)-связи между остатками фукозы на невосстанавливающем конце (рис. 3.19, слева). Однако, МС/МС спектр содержал ионы 0,3Х1 с m/z315.037 и 0,2Х1 с m/z285.029, указывающие на наличие (1^4)-связи между этими же остатками (рис. 3.19, справа). Ион очень низкой интенсивности 0,2А3 с m/z475.112 и вышеупомянутый 0,2Х0 свидетельствовали о вероятности (1^3)-связи между остатками фукозы, находящимися на восстанавливающем конце. Таким образом, методом МС/МС обнаружены фрагменты фукоидана, построенные из сульфатированных остатков фукозы с чередующимися (1-»3)- и (1->4)-связями, что также находится в соответствии с результатами предыдущих исследований (Kusaykin et al., 2006). Анализ более протяженных сульфатированных фукоолигосахаридов не дал столь четких результатов из-за большого количества шума в спектре и сложности в интерпретации.
Использование тандемной масс-спектрометрии позволило установить структурное значение в фукоидане из F. evanescensеще одного минорного компонента - галактозы. МС/МС спектр иона с m/z259.018, соответствующего [GalS03Na-Na]", давал информацию о С2 и С4/С6 сульфатировании
85
хЮ 3.4-1
3,2^
3 I 2,8^
2,6^
^2,4
8 2,2
I
1 2
ff 1.6
ii,4
ё1,2 < 1
0,8^ 0,6^ 0/N
0,2^
oJ
CH2OH ' Al CH2OH
-о//"*" Л-—ov/
Hid ,.il 1.1
MX, 0,2Y
CH2OH ,.*J CH2OH _A^
-О'' V, 7 J--------О /
■\ .J- r2/ К ОН
°>3X,
[MNa-Na]"
130 140 150 160 170 180 190 200 210 220 230 240 250 260 270 280 290 300 310 320 330 340 350 360 370 380 390 400 410 420 430 m/Z
Рис. 3.20. Тандемный ИЭР масс-спектр иона [Gal2S03Na-Na]" с m/z421.060
остатков галактозы. МС/МС спектр иона [Gal2SO Na-Na] с m/z421.060 (рис. 3.20) содержал интенсивный фрагментный ион с m/z241.000 и менее интенсивный с m/z259.012. Образование этих ионов связано с разрывом гликозидных связей и отщеплением дегидратированного остатка сульфа-тированной галактозы и сульфатированной галактозы соответственно.
Ион 0,2Х0 с m/z138.970 указывал на сульфатирование остатка галактозы на восстанавливающем конце при С2. Фрагментные ионы 0,2Х с m/z301.022 и 0,3Х1 с m/z331.032 свидетельствовали о 2-О-сульфатировании остатка галактозы, находящегося на невосстанавливающем конце. Фрагменты, характерные для сульфатирования остатка галактозы при С4, отсутствовали. Таким образом, МС/МС-анализ позволил обнаружить и установить структуру галактозосодержащих фрагментов фукоидана: Gal2S03--(l-H)-Gal и Gal-(l->4)-Gal2S03.
Галактоза входила также в состав смешанного дисахарида [Fu^Ga^SO^a-Na] с m/z405.070, присутствующего в МС-спектре. Как и в случае с предыдущими сульфатированными дисахаридами, этот ион содержал несколько изомеров, различающихся положением в них моно-сахаридных остатков и сульфатных групп. МС/МС спектр (рис. 3.21) содержал интенсивные ионы с m/z225.007 и 241.003, образование которых связано с разрывом гликозидных связей и отрывам сульфатированных остатков дегидратированной фукозы и галактозы. Фрагментные ионы 0,2Х-типа с m/z285.029 и 301.021 указывали на 2-О-сульфатирование невосстанавливающих остатков фукозы и галактозы.
Фрагментный ион оаА с m/z198.992 (на рисунке не отмечен) указывал на 4-О-сульфатирование невосстанавливающего остатка галактозы,
86
Рис. 3.21. Тандемный ИЭР масс-спектр иона [FuCjGaljSC^Na-Na]- с m/z405.070
следов 4-О-сульфатирования остатков фукозы не обнаружено. Присутствие ионов 0,2А2 с m/z345.047 и 0,2Х0, указывает на наличие (1^4)-связи, однако наличие (1-*3) -связи не может быть исключено, т.к. в таком случае, как уже было неоднократно отмечено, не образуются фрагментные ионы 0>2Х и 0>2А. Наличие иона 0>3Х0 с m/z168.979 также подтверждает присутствие структурного варианта Fuc-(1^4)-Gal-2-S03- в исследуемой смеси, т.к. для остатков сульфатированной галактозы вероятность фрагментации по типу 0,3Х была выше, чем для остатков фукозы (см. описание МС/МС спектра сульфатированной галактозы). Таким образом, масс-спектрометрический анализ показал наличие в смеси ионов Gal-(1^4)-Fuc-2-S03-, Gal-2-S03"-(l-H)-Fuc и незначительное количество Fuc-(1^4)-Gal-2-S03. Остаток галактозы на невосстанавливающем конце может быть сульфатирован при С4.
Картина масс-спектрометрической фрагментации иона [Fuc1Gal2S03Na--Na]
| m/z | Состав |
| 243.02 | [Fuc,S03Na-Na]" |
| 371.07 | [FucjSOjNa-HjO-NaJ |
| 389.08 | [Fuc2SO,Na-Na]" |
| 473.01 | [Fuc2(S03Na)2-H20-Na]' |
| 491.02 | [Fu^(S03Na)2-Na]" |
| 507.00 | [FuciGal,(S03Na)2-Na]" |
| 592.95 | [FuQ2(S03Na)3-Na]- |
| 608.90 | [FuclGall(S03Na)3-Na] |
| 783.12 | [FuC4(S03Na)2-Na]" |
| 885.07 | [Fuc4(S03Na)3-NaT |
| 987.01 | [Fuc4(S03Na)4-NaT |
| 1088.96 | [Fuc4(S03Na)5-NaT |
| 1177.20 | [Fuo;(S03Na)3-NaT |
400
600
800
1000
1200 m/z
Рис. 3.23. МАЛДИ масс-спектр отрицательных ионов олигосахаридов, полученных автогидролизом фукоидана из бурой водоросли F. evanescens
88
Структурные характеристики олигосахаридов были установлены с помощью МАЛДИ МС/МС. Основной компонент смеси — дисульфа-тированная фукобиоза имела преимущественно (1^4)-гликозидную связь между остатками фукозы (Anastyuk et al., 2012). Ее тандемный масс-спектр представлен на рисунке 3.24.
100J
гсбСН
0
Na03SO
°'2А2 gj
В]
о
Y,
°'2ХЛ
hso;
т- О)
I о
со
0,2Х,
В', аэ со
см
\
[M(2Na)-NaHS04-Na] I °<2Х',
т- СО h- CO СО СО
jjkb.
М*
100 150 200 250 300 350 400 m/z
Рис. 3.24. Тандемный МАЛДИ масс-спектр иона [Fuc (SO Na)2-Na]" c m/z491.0
По характеру интенсивностей фрагментных ионов масс-спектр оказался более похож на тандемный масс-спектр дисульфатированной фукобио-
зы, выделенной из фукоидана А.
P^laOsSOv'
Рис. 3.25. Тандемный МАЛДИ масс-спектр иона [Fuc,(SO,Na),-Na]- с m/z593.0
nodosum, где авторами было отмечено существенное количество (1^4)-связанных остатков a-L-Fucp. (Daniel et al., 2007) При этом, из спектра не следовало, есть ли в смеси (1^3)-связанные остатки a-L-Fucp.
Этот вопрос был снят при анализе иона трисульфатированной фукобиозы с m/z593.0. (рис. 3.25). Благодаря наличию дополнительной сульфатной группы, мы смогли наблюдать Y-ион с m/z344.9, соответствующий отщеплению дважды сульфатированного остатка фукозы с восстанавливающего конца молекулы. Поскольку в фу-
89
коидане из F. evanescensсульфатирование при СЗ остатка ct-L-Fucp не было обнаружено (Bilan et al., 2002), был сделан вывод о присутствии некоторого количества (1^3)-связанных остатков ct-L-Fucp (рис. 3.24).
Остатки гексозы (D-Gal) были обнаружены в составе ди- и трисульфа-тированных дисахаридов типа FucHex. По результатам анализа тандемно-го масс-спектра (рис. 3.26), было предложено 3 варианта строения дисаха-рида: Gal-2-OS03Na-(l->3)-Gal-2-OS03Na, Gal-2,4-

